



Beyond the cellulose: Oxygen isotope composition of plant lipids as a proxy for terrestrial water balance
Affiliations | Corresponding Author | Cite asKeywords: drought, plant lipids, soil-plant-atmosphere interactions, stable isotopes

- Share this article





-
Article views:11,376Cumulative count of HTML views and PDF downloads.
- Download Citation
- Rights & Permissions

Abstract

Figures and Tables
 Table 1 Species description. | 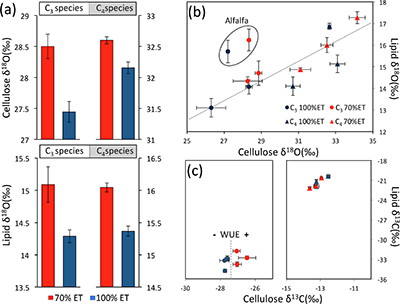 Figure 1 (a) Average δ18O values of C3 and C4 species under full evapotranspiration replenishment (100 % ET) and deficit irrigation (70 % ET). Differences between treatments were significant (t-test; P < 0.01) in all cases. (b) Relationship between cellulose and lipid δ18O values excluding alfalfa: y = 0.4449x + 1.4277; R² = 0.76 (P < 0.01). (c) Cellulose and lipid δ13C values; significant differences were observed in cellulose of C3 species (t-test; P < 0.01), but not in lipids. Error bars represent one standard deviation. | 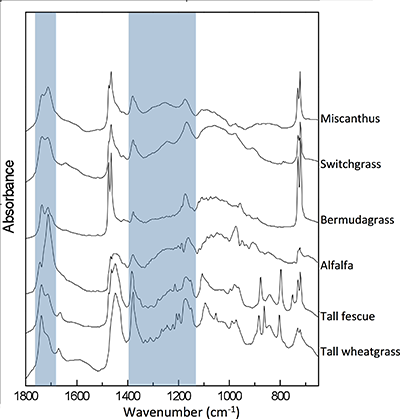 Figure 2 Difference spectra obtained by subtracting the spectra of lipid extracts of plants growing under deficit irrigation (70 % ET) from those growing under full water replenishment. Highlighted areas correspond to oxygen-containing groups (e.g., vibration from aromatic C-O, aliphatic esters and ketones). The spectra have been shifted vertically for clarity. A complete spectral characterisation is presented in Table S-3 and Figure S-1. |
| Table 1 | Figure 1 | Figure 2 |
Supplementary Figures and Tables
| Table S-1 Climate at the study location. | Table S-2 Soil properties at the study location. | Table S-3 Peaks associated with O-containing groups in the infrared spectra. | 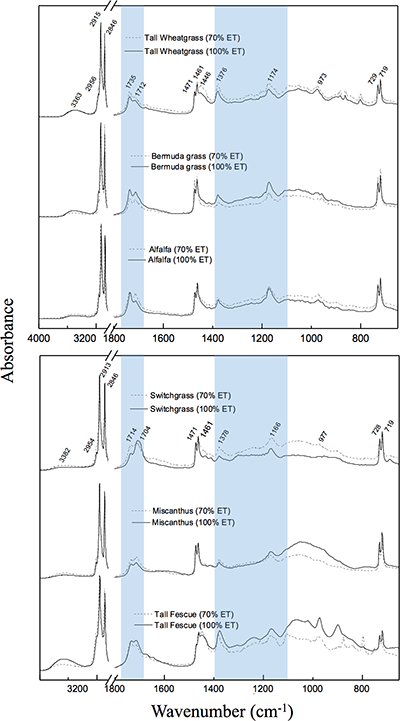 Figure S-1 Infrared spectra by species and treatment. Highlighted areas correspond to oxygen-containing groups depicted in Table S-3. Differences between full water replenished (solid lines) and deficit irrigation (dashed lines) were used to generate the drought response spectra shown in Figure 2. | 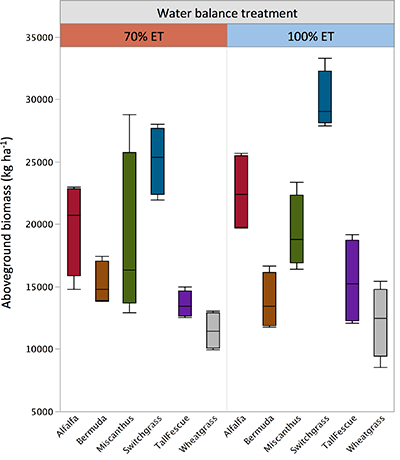 Figure S-2 Total aboveground biomass by species and treatment. Horizontal lines within the boxes represent median values. The ends of the boxes represent the 75th and 25th quantiles and whiskers span the entire data set including outliers. |
| Table S-1 | Table S-2 | Table S-3 | Figure S-1 | Figure S-2 |
top
Introduction
The study of shifting water regimes and their impact on the terrestrial ecosystems is a research frontier in which quantitative metrics are still being established. The need to characterise climate variability and to measure the impact of drought on natural and managed ecosystems has led to increasing interest in stable isotope proxies (Silva and Anand, 2013
Silva, L.C.R., Anand, M. (2013) Historical links and new frontiers in the study of forest-atmosphere interactions. Community Ecology 14, 208–218.
; Maxwell et al., 2014Maxwell, T., Silva, L.C.R., Horwath, W.R. (2014) Using multielement isotopic analysis to decipher drought impacts and adaptive management in ancient agricultural systems. Proceedings of the National Academy of Sciences of the United States of America 111, E4807–E4808.
; Silva, 2014Silva, L.C.R. (2014) Importance of climate-driven forest–savanna biome shifts in anthropological and ecological research. Proceedings of the National Academy of Sciences of the United States of America 111, E3831–E3832.
). The oxygen isotope composition (δ18O) of plant cellulose provides an integrated measure of the balance between water inputs (precipitation and/or irrigation) and losses through evapotranspiration (ET), and has been used to examine the effects of climatic variability on terrestrial systems (Kahmen et al., 2011Kahmen, A., Sachse, D., Arndt, S.K., Tu, K.P., Farrington, H., Vitousek, P.M., Dawson, T.E. (2011) Cellulose δ18O is an index of leaf-to-air vapor pressure difference (VPD) in tropical plants. Proceedings of the National Academy of Sciences of the United States of America 108, 1981–1986.
; Sternberg, 2009Sternberg, L. (2009) Oxygen stable isotope ratios of tree-ring cellulose: the next phase of understanding. New Phytologist 181, 553-562.
). Although extensively used in living plants (e.g., tree rings), the analysis of cellulose is limited by its typically short life after decomposition of plant biomass. Under particular conditions, cellulose extracted from fossil plants have been used to produce climate records that go back millions of years (Jahren and Sternberg, 2003Jahren, A.H., Sternberg, L.S.L. (2003) Humidity estimate for the middle Eocene Arctic rain forest. Geology 31, 463–466.
), but such well-preserved samples are rare. In contrast to cellulose, plant-derived lipids can persist in soils and sediments due to their low biodegradability and stabilisation into hydrophobic domains of organic matter (Matsumoto et al., 2007Matsumoto, K., Kawamura, K., Uchida, M., Shibata, Y. (2007) Radiocarbon content and stable carbon isotopic ratios of individual fatty acids in subsurface soil: Implication for selective microbial degradation and modification of soil organic matter. Geochemical Journal 41, 483–492.
; Assis et al., 2011Assis, C.P., González-Vila, F.J., Jucksch, I., González-Pérez, J.A., Neves, J.C.L., Lani, J.L., Mendonça, E.S. (2011) Lipid abundance and composition of a humic Oxisol as a function of land use. Scientia Agricola 68, 230–236.
), providing a potential long-term record of terrestrial water balance.Plant lipids may be broadly defined as hydrophobic or amphiphilic molecules originating entirely or in part from two types of biochemical subunits, ketoacyl and isoprene groups, which are building blocks for molecules of various structures and polarities, including some with no oxygen at all (e.g., alkanes in leaf surface wax) and others that carry various amount of oxygen (Fahy et al., 2009
Fahy, E., Subramaniam, S., Murphy, R.C., Nishijima, M., Raetz, C.R.H., Shimizu, T., Spener, F., van Meer, G., Wakelam, M.J.O., Dennis, E.A. (2009) Update of the LIPID MAPS comprehensive classification system for lipids. Journal of Lipid Research 50, S9–S14.
). The value of lipid hydrogen isotope analysis (δ2H) has already been recognised for specific compounds recovered from soils and sediments (Sachse et al., 2012Sachse, D., Billault, I., Bowen G.J., Chikaraishi, Y., Dawson, T.E., Feakins, S.J., Freeman, K.H., Magill, C.R., McInerney, F.A., van der Meer, M.T.J., Polissar, P., Robins, R.J., Sachs, J.P., Schmidt, H.-L., Sessions, A.L., White, J.W.C., West, J.B., Kahmen, A. (2012) Molecular Paleohydrology: Interpreting the Hydrogen-Isotopic Composition of Lipid Biomarkers from Photosynthesizing Organisms. Annual Review of Earth and Planetary Sciences 40, 221–249.
). The study of lipid δ18O signals has the potential to improve δ2H records by, for example, allowing water loss to be partitioned into evaporation and transpiration (Voelker et al., 2014Voelker, S., Brooks, J., Meinzer, F., Roden, J., Pazdur, A., Pawelczyk, S., Hartsough, P., Snyder, K., Plavcová, L., Šantrůček, J. (2014) Reconstructing relative humidity from plant δ18O and δD as deuterium deviations from the global meteoric water line. Ecological Applications 24, 960–975.
).The first step towards the use of lipid δ18O records is to demonstrate that a signal related to changes in water balance is recorded in such compounds. The second step is then to demonstrate that this signal is preserved in soils and sediments. The present study is concerned with the first step. However, without losing sight of the second step, we imposed the restriction that lipid extracts should contain only the most apolar compounds present in the plant biomass, under the assumption that these are also the most likely to persist after deposition (Matsumoto et al., 2007
Matsumoto, K., Kawamura, K., Uchida, M., Shibata, Y. (2007) Radiocarbon content and stable carbon isotopic ratios of individual fatty acids in subsurface soil: Implication for selective microbial degradation and modification of soil organic matter. Geochemical Journal 41, 483–492.
). To advance the interpretation of lipid δ18O signals, we compared measurements of cellulose and hexane-extractable compounds of C3 and C4 species, grown under contrasting water regimes in replicated field experiments. In addition, we analysed the carbon isotope composition (δ13C) of cellulose and lipids as a way to assess plant water-use efficiency (Farquhar and Von Caemmerer, 1982Farquhar, G.D., Von Caemmerer, S. (1982) Modelling of photosynthetic response to environmental conditions. In: Lange, O.L., Nobel, P.S., Osmond, C.B., Ziegler, H. (Eds.) Encyclopedia of Plant Physiology 549–587.
). Alongside these isotopic measurements, we used spectroscopic analyses to qualitatively describe the molecular composition of lipid extracts, and all results are integrated into a consideration of potential applications in ancient and contemporary settings.top
Methods
Field Experiment. The field experiment was conducted at the University of California, Five Points Experimental Station (36°20'10.28"N, 120° 6'38.40"W). This site experiences hot summers, cool and dry winters, and average annual precipitation of 173 mm. Plots of 27 m2 were established under homogeneous edaphic conditions (Tables S-1 and S-2) in which three C3 and three C4 species were grown (Table 1). Following a complete randomised block design with four field replications, monocultures of each species were established using sprinkler irrigation (2008), later converted to flood irrigation (2009), after which two irrigation treatments were imposed through 2011. Daily potential ET was obtained from the on-site meteorological station and multiplied by species-specific coefficients (Allen and Pruitt, 1991
Allen, R.G., Pruitt, W.O. (1991) FAO‐24 Reference Evapotranspiration Factors. Journal of Irrigation and Drainage Engineering 117, 758–773.
). The amount of water to be applied was determined based on 100 % or 70 % of the potential ET, representing full water loss replenishment and deficit irrigation (imposed drought) treatments, respectively. Plots were irrigated when the accumulated ET reached 150 mm, for an average of 11 irrigation events per year. Precipitation water was factored into the calculation for irrigation. The average annual δ18O of the precipitation is about -7.8 ‰. Irrigation water was a combination of aqueduct source (originating in the Sacramento-San Joaquin Delta region) and a groundwater well at a depth of 150-180 metres. The same irrigation water was used in all plots throughout the experiment.Table 1 Species description.
| Species | Scientific name | Variety | Metabolim |
| Miscanthus | Miscanthus giganteus | Hybrid giganteus | Warm-season C4 |
| Switchgrass | Panicum virgatum | Alamo | Warm-season C4 |
| Bermudagrass | Cynodon dactylon | Giant NK 37 | Warm-season C4 |
| Alfalfa | Medicago sativa | Big Kaw | Warm-season C3 |
| Tall Fescue | Festuca arundinacea | Fawn | Cool-season C3 |
| Tall wheatgrass | Agropyron elongatum | Jose | Cool-season C3 |
Sampling and analysis. At the end of the experiment, the aboveground biomass of all plots was harvested and ground to <0.5 mm for analysis. Pure alpha cellulose was isolated (Brendel et al., 2000
Brendel, O., Iannetta, P.P.M., Stewart, D. (2000) A rapid and simple method to isolate pure alpha-cellulose. Phytochemical Analysis 11, 7–10.
) and the lipid fraction was obtained by Soxhlet extraction with hexane for 10 hours. Although less efficient at extracting total lipids, as compared to more polar solvent mixtures (e.g., chloroform:methanol), hexane was chosen to minimise extraction of transient polar lipids such as phospholipids and degradation products of chlorophyll. The isotopic composition of oxygen was determined using an Elementar PyroCube (Elementar Analysensysteme, Hanau, Germany) interfaced to a PDZ Europa 20-20 IRMS (Sercon Ltd., Cheshire, UK). Three analytical replicates were used for each plot and working standards of cellulose and hexadecyl palmitate were interspersed throughout each run to determine analytical precision. Carbon isotopic composition of cellulose and lipids was determined using a PDZ Europa ANCA-GSL elemental analyser interfaced to a PDZ Europa 20-20 IRMS (Sercon Ltd., Cheshire, UK). A certified analytical standard of known δ13C was used to confirm accuracy and precision. Data are expressed as δ18O and δ13C (‰) relative to Vienna Standard Mean Ocean Water (VSMOW) and Pee Dee Belemnite (VPDB), respectively.Elemental analysis of hexane-extracted material showed an oxygen content of 6-11 % and a carbon content of 71-79 % (w/w). Fourier Transform Infrared Spectroscopy (FTIR) was used to describe the molecular composition of hexane extracts, focusing on oxygen-containing groups. Samples were analysed using attenuated total reflectance (ATR) FTIR. The spectra were collected on a Thermo Nicolet 6700 spectrophotometer (Thermo Scientific, Madison, WI) with a diamond single bounce accessory (GladiATR, PIKE Technologies). After collection, the spectra were normalised (to the aliphatic C-H peak at approximately 2913 cm-1) prior to performing spectral subtraction. All FTIR spectra were collected in triplicate using 4 cm-1 resolution and 1.2 kHz scanning speed for a total of 128 co-added scans per analysis (Parikh et al., 2014
Parikh, S. J., Mukome, F.N.D., Zhang, X. (2014) ATR–FTIR spectroscopic evidence for biomolecular phosphorus and carboxyl groups facilitating bacterial adhesion to iron oxides. Colloids and Surfaces Biointerfaces 119, 38–46.
).Statistical analysis. Average differences in isotopic composition were compared using paired t-tests. When appropriate, least-square regressions were performed to evaluate the relationship between cellulose and lipid isotopic ratios. Average data points and standard deviations were plotted for each species, blocked by water balance treatment, incorporating 4 field replicates each. Molecular characterisation of lipids was performed separately for each species and treatment.
top
Results and Discussion
Isotopic composition of cellulose and lipids. Irrigation treatments caused clear changes in lipid and cellulose δ18O values. Despite significant differences among species, plants growing under drought (70 % ET) were always enriched in 18O relative to plants growing under full ET replenishment (Fig. 1a). In cellulose, 18O enrichment in response to drought was clearer in C3 (>1 ‰) compared to C4 (~0.5 ‰) plants, while in lipids, consistent responses were observed (~0.75 ‰) with no statistical difference between C3 and C4 plants. We found a strong linear relationship between cellulose and lipid δ18O values (P < 0.01), reflecting consistent responses across species (Fig. 1b).
The effect of drought was also evident in cellulose δ13C, reflecting expected differences between plant metabolism (Pearcy and Ehleringer, 1984
Pearcy, R.W., Ehleringer, J. (1984) Comparative ecophysiology of C3 and C4 plants. Plant, Cell and Environment 7, 1–13.
), and showing a clear separation in response to water balance treatments in C3 plants (Fig. 1c). This separation confirms an anticipated increase in water-use efficiency under drought, which was surprisingly not apparent in lipid δ13C values, and may be explained by the heterogeneous composition of lipid extracts (discussed below). While 13C fractionation in C3 plants is controlled by the stomata-regulated ratio of intercellular to ambient CO2 concentrations (ci/ca), variation in C4 bundle-sheath leakiness can either dampen or amplify the effects of ci/ca, so that δ13C measurements cannot be used to assess water-use efficiency in C4 plants (Cernusak et al., 2013Cernusak, L.A., Ubierna, N., Winter, K., Holtum, J.A.M., Marshall, J.D., Farquhar, G.D. (2013) Environmental and physiological determinants of carbon isotope discrimination in terrestrial plants. The New Phytologist 200, 950–965.
). Nevertheless, we found 18O enrichment of similar magnitude and direction in C3 and C4 lipids under drought, demonstrating the value of δ18O measurements as a general proxy for water balance.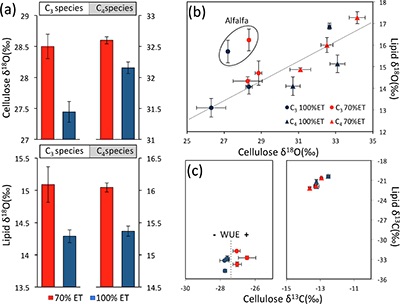
Figure 1 (a) Average δ18O values of C3 and C4 species under full evapotranspiration replenishment (100 % ET) and deficit irrigation (70 % ET). Differences between treatments were significant (t-test; P < 0.01) in all cases. (b) Relationship between cellulose and lipid δ18O values excluding alfalfa: y = 0.4449x + 1.4277; R² = 0.76 (P < 0.01). (c) Cellulose and lipid δ13C values; significant differences were observed in cellulose of C3 species (t-test; P < 0.01), but not in lipids. Error bars represent one standard deviation.
Interpreting differences in isotopic composition. Leaf and xylem water are both sources of oxygen incorporated into organic compounds during biosynthesis, but each of these sources contributes a distinct isotopic signal (Sternberg, 2009
Sternberg, L. (2009) Oxygen stable isotope ratios of tree-ring cellulose: the next phase of understanding. New Phytologist 181, 553-562.
). Reductions in plant water supply are expected to increases the contribution of isotopically enriched leaf water, while decreasing the flux of relatively unenriched xylem water to the leaf (Ogée et al., 2007Ogée, J., Cuntz, M., Peylin, P., Bariac, T. (2007) Non-steady-state, non-uniform transpiration rate and leaf anatomy effects on the progressive stable isotope enrichment of leaf water along monocot leaves. Plant, Cell and Environment 30, 367–387.
). This process is influenced by the distribution of leaf veins, which caused the greatest 18O enrichment in lipids relative to cellulose in alfalfa, the only dicot species. This observation is consistent with previous work on leaf δ2H values showing significant differences in 2H enrichment of the n-C29 alkane biomarker among plant growth forms, in which dicot species were the most and graminoids the least enriched in 2H, respectively (Sachse et al., 2012Sachse, D., Billault, I., Bowen G.J., Chikaraishi, Y., Dawson, T.E., Feakins, S.J., Freeman, K.H., Magill, C.R., McInerney, F.A., van der Meer, M.T.J., Polissar, P., Robins, R.J., Sachs, J.P., Schmidt, H.-L., Sessions, A.L., White, J.W.C., West, J.B., Kahmen, A. (2012) Molecular Paleohydrology: Interpreting the Hydrogen-Isotopic Composition of Lipid Biomarkers from Photosynthesizing Organisms. Annual Review of Earth and Planetary Sciences 40, 221–249.
).Fatty acid transfer reactions within the plastid yield a significant reduction in 18O content of water in relation to the cytosol (Pollard and Ohlrogge, 1999
Pollard, M., Ohlrogge, J. (1999) Testing models of fatty acid transfer and lipid synthesis in spinach leaf using in vivo oxygen-18 labeling. Plant Physiology 121, 1217–1226.
) contributing to differences between the isotopic composition of lipids and cellulose. It is important to note that lipids contain far less oxygen than cellulose and do not undergo the postsynthetic isotopic exchange that happens during the hydration of carbonyl groups of the intermediates of cellulose synthesis (Sternberg and DeNiro, 1983Sternberg, L., DeNiro, M.J.D. (1983) Biogeochemical implications of the isotopic equilibrium fractionation factor between the oxygen atoms of acetone and water. Geochimica et Cosmochimica Acta 47, 2271–2274.
). Therefore, intrinsic species traits and biosynthetic differences explain the offset between lipid and cellulose δ18O values across species (Fig. 1b).Our observations show that the balance between water input and evapotranspiration can be directly interpreted from lipid oxygen isotopes in monocultures of C3 and C4 species, providing a powerful tool for selecting traits that would optimise water-use efficiency under field conditions. However, the large variation in 18O enrichment between species indicates that the effect of a diverse vegetation cover could confound water balance signals. Therefore, analyses of lipids in natural ecosystems are more likely to successfully reconstruct climate variability where vegetation composition is known to be stable. This could be accomplished by appropriate evaluation of vegetation composition in contemporary settings, and palaeoecological reconstructions in ancient settings, based on companion proxies for vegetation change (e.g., fossil pollen and δ13C of organic matter; Silva and Anand, 2013
Silva, L.C.R., Anand, M. (2013) Historical links and new frontiers in the study of forest-atmosphere interactions. Community Ecology 14, 208–218.
; Silva, 2014Silva, L.C.R. (2014) Importance of climate-driven forest–savanna biome shifts in anthropological and ecological research. Proceedings of the National Academy of Sciences of the United States of America 111, E3831–E3832.
).Characterisation of lipid extracts. The FTIR spectra of lipid extracts show that substantial differences in molecular composition exist among species and in response to growing conditions. The difference spectra (Fig. 2), obtained by subtracting results of water-limited from non-limited fields, show that all common oxygen-containing functional groups found in lipids are more abundant under drought. These groups include long-chain aliphatic esters, ketones, carboxylic acids, ethers, alcohols, as well as aromatic compounds (Table S-3 and Fig. S-1). This observation is consistent with general variations in plant lipid composition (Fahy et al., 2009
Fahy, E., Subramaniam, S., Murphy, R.C., Nishijima, M., Raetz, C.R.H., Shimizu, T., Spener, F., van Meer, G., Wakelam, M.J.O., Dennis, E.A. (2009) Update of the LIPID MAPS comprehensive classification system for lipids. Journal of Lipid Research 50, S9–S14.
), as drought stress tends to decrease the abundance of more polar lipids (e.g., phospholipids) while increasing the relative abundance of nonpolar compounds (Harwood, 1998Harwood, J. (1998) Plant Lipid Biosynthesis: Fundamentals and Agricultural Applications 378. Society for Experimental Biology, Seminar Series 67, Cambridge University Press, 378 pp.
).Since the 1970s, when the use of δ18O values obtained from tree rings was first established, cellulose δ18O records have been used as a proxy for drought stress. Classic conceptual models (Scheidegger et al., 2000
Scheidegger, Y., Saurer, M., Bahn, M., Siegwolf, R. (2000) Linking stable oxygen and carbon isotopes with stomatal conductance and photosynthetic capacity: a conceptual model. Oecologia 125, 350–357.
) have coupled δ18O and δ13C values to distinguish differences in stomatal response from photosynthetic capacity. In the present dataset, this link was apparent in cellulose but not in lipids, probably due to the heterogeneity of bulk lipid extracts. Nevertheless, lipid extracts of C3 and C4 plants showed clear isotopic signals in response to drought. This result demonstrates that δ18O values of bulk plant lipids serve as a more general proxy for water balance than δ13C records, which are limited to C3 plants and therefore necessarily exclude many important crops and tropical ecosystems. This is particularly important because shifts in δ18O signals in response to treatments were independent of variation in plant productivity (Fig. S-2) and large interspecific differences in molecular composition of hexane extracts (Fig. S-1).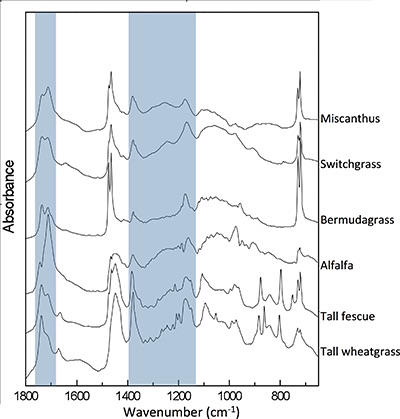
Figure 2 Difference spectra obtained by subtracting the spectra of lipid extracts of plants growing under deficit irrigation (70 % ET) from those growing under full water replenishment. Highlighted areas correspond to oxygen-containing groups (e.g., vibration from aromatic C-O, aliphatic esters and ketones). The spectra have been shifted vertically for clarity. A complete spectral characterisation is presented in Table S-3 and Figure S-1.
The value of a new lipid proxy. The present study provides empirical evidence to support the use of lipid oxygen isotopic composition as a proxy for water balance in contemporary settings, which also has the potential to improve reconstructions of past climates. Measurements of hydrogen isotopes in plant lipids preserved in soils and sediments have been previously used to reconstruct climate variability, but these isotopic signals are sensitive to biochemical, physiological, and environmental influences (Sachse et al., 2012
Sachse, D., Billault, I., Bowen G.J., Chikaraishi, Y., Dawson, T.E., Feakins, S.J., Freeman, K.H., Magill, C.R., McInerney, F.A., van der Meer, M.T.J., Polissar, P., Robins, R.J., Sachs, J.P., Schmidt, H.-L., Sessions, A.L., White, J.W.C., West, J.B., Kahmen, A. (2012) Molecular Paleohydrology: Interpreting the Hydrogen-Isotopic Composition of Lipid Biomarkers from Photosynthesizing Organisms. Annual Review of Earth and Planetary Sciences 40, 221–249.
), which can be better constrained by dual isotope analyses (Scheidegger et al., 2000Scheidegger, Y., Saurer, M., Bahn, M., Siegwolf, R. (2000) Linking stable oxygen and carbon isotopes with stomatal conductance and photosynthetic capacity: a conceptual model. Oecologia 125, 350–357.
; Maxwell et al., 2014Maxwell, T., Silva, L.C.R., Horwath, W.R. (2014) Using multielement isotopic analysis to decipher drought impacts and adaptive management in ancient agricultural systems. Proceedings of the National Academy of Sciences of the United States of America 111, E4807–E4808.
; Voelker et al., 2014Voelker, S., Brooks, J., Meinzer, F., Roden, J., Pazdur, A., Pawelczyk, S., Hartsough, P., Snyder, K., Plavcová, L., Šantrůček, J. (2014) Reconstructing relative humidity from plant δ18O and δD as deuterium deviations from the global meteoric water line. Ecological Applications 24, 960–975.
). The much lower abundance of oxygen relative to hydrogen atoms in plant lipids has so far limited the production of δ18O records from specific compounds. However, persistent oxygen-containing lipids, such as long-chain n-alkanoic acids, are commonly recovered in sedimentary profiles and analysed for δ2H (Feakins et al., 2007Feakins, S.J., Eglinton, T.I., de Menocal, P.B. (2007) A comparison of biomarker records of northeast African vegetation from lacustrine and marine sediments (ca. 3.40 Ma). Organic Geochemistry 38, 1607–1624.
), and could lead to water balance reconstructions with improved precision. In the case of hydrogen, the key to success has been the study of specific compounds in which the effects of hydrological shifts and biosynthesis can be isolated. Similarly, the analysis of oxygen isotopes of hemicellulose degradation products in soil has been recently shown to reflect the balance between precipitation and evapotranspiration across broad latitudinal gradients (Tuthorn et al., 2014Tuthorn, M., Zech, M., Ruppenthal, M., Oelmann, Y., Kahmen, A., del Valle, H.F., Wilcke, W., Glaser, B. (2014) Oxygen isotope ratios (18O/16O) of hemicellulose-derived sugar biomarkers in plants, soils and sediments as paleoclimate proxy II: Insight from a climate transect study. Geochimica et Cosmochimica Acta 126, 624–634.
; Zech et al., 2014Zech, M., Mayr, C., Tuthorn, M., Leiber-Sauheitl, K., Glaser, B. (2014) Oxygen isotope ratios (18O/16O) of hemicellulose-derived sugar biomarkers in plants, soils and sediments as paleoclimate proxy I: Insight from a climate chamber experiment. Geochimica et Cosmochimica Acta 126, 614–623.
). We expect bulk lipid samples to serve as a basis from which compound- specific oxygen and hydrogen records can be developed to better understand present and decipher past impacts of climatic change on terrestrial systems.top
Acknowledgements
We thank Johan Six, Bruce Linquist and Chris Van Kessel for logistic support in conducting the field experiments; Sanjai Parikh for laboratory facilities and equipment necessary for the infrared spectral analysis; Leonel Sternberg and participants of the Goldschmidt Conference 2014, session on Plant Compounds as Ecohydrological Tracers, for valuable discussion. This work is supported by the California Department of Food and Agriculture Project #26491 (LS), by the NSF-NIFA Water Sustainability and Climate Program #2014-67003-22077 (WH; LS), and the J. G. Boswell Endowed Chair in Soil Science.
Editor: Liane G. Benning
top
References
Allen, R.G., Pruitt, W.O. (1991) FAO‐24 Reference Evapotranspiration Factors. Journal of Irrigation and Drainage Engineering 117, 758–773.
 Show in context
Show in context Daily potential ET was obtained from the on-site meteorological station and multiplied by species-specific coefficients (Allen and Pruitt, 1991).
View in article
Assis, C.P., González-Vila, F.J., Jucksch, I., González-Pérez, J.A., Neves, J.C.L., Lani, J.L., Mendonça, E.S. (2011) Lipid abundance and composition of a humic Oxisol as a function of land use. Scientia Agricola 68, 230–236.
Show in context In contrast to cellulose, plant-derived lipids can persist in soils and sediments due to their low biodegradability and stabilisation into hydrophobic domains of organic matter (Matsumoto et al., 2007; Assis et al., 2011), providing a potential long-term record of terrestrial water balance.
View in article
Brendel, O., Iannetta, P.P.M., Stewart, D. (2000) A rapid and simple method to isolate pure alpha-cellulose. Phytochemical Analysis 11, 7–10.
Show in context Pure alpha cellulose was isolated (Brendel et al., 2000) and the lipid fraction was obtained by Soxhlet extraction with hexane for 10 hours.
View in article
Cernusak, L.A., Ubierna, N., Winter, K., Holtum, J.A.M., Marshall, J.D., Farquhar, G.D. (2013) Environmental and physiological determinants of carbon isotope discrimination in terrestrial plants. The New Phytologist 200, 950–965.
Show in context While 13C fractionation in C3 plants is controlled by the stomata-regulated ratio of intercellular to ambient CO2 concentrations (ci/ca), variation in C4 bundle-sheath leakiness can either dampen or amplify the effects of ci/ca, so that δ13C measurements cannot be used to assess water-use efficiency in C4 plants (Cernusak et al., 2013).
View in article
Fahy, E., Subramaniam, S., Murphy, R.C., Nishijima, M., Raetz, C.R.H., Shimizu, T., Spener, F., van Meer, G., Wakelam, M.J.O., Dennis, E.A. (2009) Update of the LIPID MAPS comprehensive classification system for lipids. Journal of Lipid Research 50, S9–S14.
Show in context Plant lipids may be broadly defined as hydrophobic or amphiphilic molecules originating entirely or in part from two types of biochemical subunits, ketoacyl and isoprene groups, which are building blocks for molecules of various structures and polarities, including some with no oxygen at all (e.g., alkanes in leaf surface wax) and others that carry various amount of oxygen (Fahy et al., 2009).
View in article
This observation is consistent with general variations in plant lipid composition (Fahy et al., 2009), as drought stress tends to decrease the abundance of more polar lipids (e.g., phospholipids) while increasing the relative abundance of nonpolar compounds (Harwood, 1998).
View in article
Farquhar, G.D., Von Caemmerer, S. (1982) Modelling of photosynthetic response to environmental conditions. In: Lange, O.L., Nobel, P.S., Osmond, C.B., Ziegler, H. (Eds.) Encyclopedia of Plant Physiology 549–587.
Show in context In addition, we analysed the carbon isotope composition (δ13C) of cellulose and lipids as a way to assess plant water-use efficiency (Farquhar and Von Caemmerer, 1982).
View in article
Feakins, S.J., Eglinton, T.I., de Menocal, P.B. (2007) A comparison of biomarker records of northeast African vegetation from lacustrine and marine sediments (ca. 3.40 Ma). Organic Geochemistry 38, 1607–1624.
Show in context However, persistent oxygen-containing lipids, such as long-chain n-alkanoic acids, are commonly recovered in sedimentary profiles and analysed for δ2H (Feakins et al., 2007), and could lead to climate reconstructions with improved precision.
View in article
Harwood, J. (1998) Plant Lipid Biosynthesis: Fundamentals and Agricultural Applications 378. Society for Experimental Biology, Seminar Series 67, Cambridge University Press, 378 pp.
Show in context This observation is consistent with general variations in plant lipid composition (Fahy et al., 2009), as drought stress tends to decrease the abundance of more polar lipids (e.g., phospholipids) while increasing the relative abundance of nonpolar compounds (Harwood, 1998).
View in article
Jahren, A.H., Sternberg, L.S.L. (2003) Humidity estimate for the middle Eocene Arctic rain forest. Geology 31, 463–466.
Show in context Under particular conditions, cellulose extracted from fossil plants have been used to produce climate records that go back millions of years (Jahren and Sternberg, 2003), but such well-preserved samples are rare.
View in article
Kahmen, A., Sachse, D., Arndt, S.K., Tu, K.P., Farrington, H., Vitousek, P.M., Dawson, T.E. (2011) Cellulose δ18O is an index of leaf-to-air vapor pressure difference (VPD) in tropical plants. Proceedings of the National Academy of Sciences of the United States of America 108, 1981–1986.
Show in context Measurements of oxygen isotopes (δ18O) of plant cellulose provide an integrated measure of the balance between water input (precipitation and/or irrigation) and losses through evapotranspiration (ET), and have been used to examine the effects of climatic variability (Kahmen et al., 2011; Sternberg, 2009).
View in article
Matsumoto, K., Kawamura, K., Uchida, M., Shibata, Y. (2007) Radiocarbon content and stable carbon isotopic ratios of individual fatty acids in subsurface soil: Implication for selective microbial degradation and modification of soil organic matter. Geochemical Journal 41, 483–492.
Show in context In contrast to cellulose, plant-derived lipids can persist in soils and sediments due to their low biodegradability and stabilisation into hydrophobic domains of organic matter (Matsumoto et al., 2007; Assis et al., 2011), providing a potential long-term record of terrestrial water balance.
View in article
However, without losing sight of the second step, we imposed the restriction that lipid extracts should contain only the most apolar compounds present in the plant biomass, under the assumption that these are also the most likely to persist after deposition (Matsumoto et al., 2007).
View in article
Maxwell, T., Silva, L.C.R., Horwath, W.R. (2014) Using multielement isotopic analysis to decipher drought impacts and adaptive management in ancient agricultural systems. Proceedings of the National Academy of Sciences of the United States of America 111, E4807–E4808.
Show in context The need to characterise climate variability and to measure the impact of drought on natural and managed ecosystems has led to increasing interest in stable isotope proxies (Silva and Anand, 2013; Maxwell et al., 2014; Silva, 2014).
View in article
Measurements of hydrogen isotopes in plant lipids preserved in soils and sediments have been previously used to reconstruct climate variability, but these isotopic signals are sensitive to biochemical, physiological, and environmental influences (Sachse et al., 2012), which can be better constrained by dual isotope analyses (Scheidegger et al., 2000; Maxwell et al., 2014; Voelker et al., 2014).
View in article
Ogée, J., Cuntz, M., Peylin, P., Bariac, T. (2007) Non-steady-state, non-uniform transpiration rate and leaf anatomy effects on the progressive stable isotope enrichment of leaf water along monocot leaves. Plant, Cell and Environment 30, 367–387.
Show in context Reductions in plant water supply are expected to increases the contribution of isotopically enriched leaf water, while decreasing the flux of unenriched xylem water to the leaf (Ogée et al., 2007).
View in article
Parikh, S. J., Mukome, F.N.D., Zhang, X. (2014) ATR–FTIR spectroscopic evidence for biomolecular phosphorus and carboxyl groups facilitating bacterial adhesion to iron oxides. Colloids and Surfaces Biointerfaces 119, 38–46.
Show in context All FTIR spectra were collected in triplicate using 4 cm-1 resolution and 1.2 kHz scanning speed for a total of 128 co-added scans per analysis (Parikh et al., 2014).
View in article
Pearcy, R.W., Ehleringer, J. (1984) Comparative ecophysiology of C3 and C4 plants. Plant, Cell and Environment 7, 1–13.
Show in context The effect of drought was also evident in cellulose δ13C, reflecting expected differences between metabolism (Pearcy and Ehleringer, 1984), and showing a clear separation in response to treatments in C3 plants (Fig. 1c).
View in article
Pollard, M., Ohlrogge, J. (1999) Testing models of fatty acid transfer and lipid synthesis in spinach leaf using in vivo oxygen-18 labeling. Plant Physiology 121, 1217–1226.
Show in context Additionally, fatty acid transfer reactions within the plastid yield a significant reduction in 18O content of water in relation to the cytosol (Pollard and Ohlrogge, 1999) contributing to differences between the isotopic composition of lipids and cellulose.
View in article
Sachse, D., Billault, I., Bowen G.J., Chikaraishi, Y., Dawson, T.E., Feakins, S.J., Freeman, K.H., Magill, C.R., McInerney, F.A., van der Meer, M.T.J., Polissar, P., Robins, R.J., Sachs, J.P., Schmidt, H.-L., Sessions, A.L., White, J.W.C., West, J.B., Kahmen, A. (2012) Molecular Paleohydrology: Interpreting the Hydrogen-Isotopic Composition of Lipid Biomarkers from Photosynthesizing Organisms. Annual Review of Earth and Planetary Sciences 40, 221–249.
Show in context The value of lipid hydrogen isotope analysis (δ2H) has already been recognised for specific compounds recovered from soils and sediments (Sachse et al., 2012).
View in article
This observation is consistent with previous work on leaf δ2H values showing significant differences in 2H enrichment of the n-C29 alkane biomarker among plant growth forms, in which dicot species were the most and graminoids the least enriched in 2H, respectively (Sachse et al., 2012).
View in article
Measurements of hydrogen isotopes in plant lipids preserved in soils and sediments have been previously used to reconstruct climate variability, but these isotopic signals are sensitive to biochemical, physiological, and environmental influences (Sachse et al., 2012), which can be better constrained by dual isotope analyses (Scheidegger et al., 2000; Maxwell et al., 2014; Voelker et al., 2014).
View in article
Scheidegger, Y., Saurer, M., Bahn, M., Siegwolf, R. (2000) Linking stable oxygen and carbon isotopes with stomatal conductance and photosynthetic capacity: a conceptual model. Oecologia 125, 350–357.
Show in context Previous conceptual models (Scheidegger et al., 2000) have proposed linking δ18O and δ13C values to distinguish differences in stomatal response from photosynthetic capacity.
View in article
Measurements of hydrogen isotopes in plant lipids preserved in soils and sediments have been previously used to reconstruct climate variability, but these isotopic signals are sensitive to biochemical, physiological, and environmental influences (Sachse et al., 2012), which can be better constrained by dual isotope analyses (Scheidegger et al., 2000; Maxwell et al., 2014; Voelker et al., 2014).
View in article
Silva, L.C.R., Anand, M. (2013) Historical links and new frontiers in the study of forest-atmosphere interactions. Community Ecology 14, 208–218.
Show in context The need to characterise climate variability and to measure the impact of drought on natural and managed ecosystems has led to increasing interest in stable isotope proxies (Silva and Anand, 2013; Maxwell et al., 2014; Silva, 2014).
View in article
This could be accomplished by appropriate evaluation of vegetation composition in contemporary settings, and palaeoecological reconstructions in ancient settings, based on companion proxies (e.g., fossil pollen and δ13C of organic matter; Silva and Anand, 2013; Silva, 2014).
View in article
Silva, L.C.R. (2014) Importance of climate-driven forest–savanna biome shifts in anthropological and ecological research. Proceedings of the National Academy of Sciences of the United States of America 111, E3831–E3832.
Show in context The need to characterise climate variability and to measure the impact of drought on natural and managed ecosystems has led to increasing interest in stable isotope proxies (Silva and Anand, 2013; Maxwell et al., 2014; Silva, 2014).
View in article
This could be accomplished by appropriate evaluation of vegetation composition in contemporary settings, and palaeoecological reconstructions in ancient settings, based on companion proxies (e.g., fossil pollen and δ13C of organic matter; Silva and Anand, 2013; Silva, 2014).
View in article
Sternberg, L. (2009) Oxygen stable isotope ratios of tree-ring cellulose: the next phase of understanding. New Phytologist 181, 553-562.
Show in context Measurements of oxygen isotopes (δ18O) of plant cellulose provide an integrated measure of the balance between water input (precipitation and/or irrigation) and losses through evapotranspiration (ET), and have been used to examine the effects of climatic variability (Kahmen et al., 2011; Sternberg, 2009).
View in article
Leaf and xylem water are both sources of oxygen incorporated into organic compounds during photosynthesis, but each of these sources contributes a distinct isotopic signal (Sternberg, 2009).
View in article
Sternberg, L., DeNiro, M.J.D. (1983) Biogeochemical implications of the isotopic equilibrium fractionation factor between the oxygen atoms of acetone and water. Geochimica et Cosmochimica Acta 47, 2271–2274.
Show in context It is important to note that lipids contain far less oxygen than cellulose and do not undergo the isotopic exchange between ketone-oxygen and unenriched water that happens during cellulose synthesis (Sternberg and DeNiro, 1983).
View in article
Tuthorn, M., Zech, M., Ruppenthal, M., Oelmann, Y., Kahmen, A., del Valle, H.F., Wilcke, W., Glaser, B. (2014) Oxygen isotope ratios (18O/16O) of hemicellulose-derived sugar biomarkers in plants, soils and sediments as paleoclimate proxy II: Insight from a climate transect study. Geochimica et Cosmochimica Acta 126, 624–634.
Show in context Similarly, the analysis of oxygen isotopes of hemicellulose degradation products in soil has been recently shown to record the balance between precipitation and evapotranspiration across broad latitudinal gradients (Tuthorn et al., 2014; Zech et al., 2014).
View in article
Voelker, S., Brooks, J., Meinzer, F., Roden, J., Pazdur, A., Pawelczyk, S., Hartsough, P., Snyder, K., Plavcová, L., Šantrůček, J. (2014) Reconstructing relative humidity from plant δ18O and δD as deuterium deviations from the global meteoric water line. Ecological Applications 24, 960–975.
Show in context The study of lipid δ18O signals has the potential to improve δ2H records by, for example, allowing effects to be partitioned into evaporation and transpiration (Voelker et al., 2014).
View in article
Measurements of hydrogen isotopes in plant lipids preserved in soils and sediments have been previously used to reconstruct climate variability, but these isotopic signals are sensitive to biochemical, physiological, and environmental influences (Sachse et al., 2012), which can be better constrained by dual isotope analyses (Scheidegger et al., 2000; Maxwell et al., 2014; Voelker et al., 2014).
View in article
Zech, M., Mayr, C., Tuthorn, M., Leiber-Sauheitl, K., Glaser, B. (2014) Oxygen isotope ratios (18O/16O) of hemicellulose-derived sugar biomarkers in plants, soils and sediments as paleoclimate proxy I: Insight from a climate chamber experiment. Geochimica et Cosmochimica Acta 126, 614–623.
Show in context Similarly, the analysis of oxygen isotopes of hemicellulose degradation products in soil has been recently shown to record the balance between precipitation and evapotranspiration across broad latitudinal gradients (Tuthorn et al., 2014; Zech et al., 2014).
View in article
top
Supplementary Information
Table S-1 Climate at the study location.
| Climatic characteristics | Five Points |
| Köppen climate classification | (BSk) Semi-arid steppe |
| Average annual temp. (°C) | 17.1 |
| Average max. annual temp. (°C) | 25 |
| Average min. annual temp. (°C) | 9.2 |
| Average annual precipitation (mm) | 173 |
| Frost free days | 335 |
| Altitude (m) | 70 |
Table S-2 Soil properties at the study location.
| Soil Properties | |
| Clay (g kg-1) | 310 |
| Silt (g kg-1) | 340 |
| Sand (g kg-1) | 350 |
| pH (saturated paste extract) | 7.6 |
| CEC (cmolc kg-1) | 30.7 |
| Olsen-P (mg kg-1) | 7.4 |
| NO3-N (mg kg-1) | 10,6 |
| Extractable K (mg kg-1) | 439 |
| Organic matter (g kg-1) | 9.5 |
| Total N (g kg-1) | 0.7 |
Table S-3 Peaks associated with O-containing groups in the infrared spectra.
| Wavenumber (cm-1) | Assignment | Reference |
| 3360 | ν(O-H) vibrations in OH | Dubis et al., 1999 |
| 2950 | ν(C-H) asymmetric vibrations in lipid CH3 | Wallach et al., 1979; Casal and Mantsch, 1984 |
| 2915 | ν(C-H) asymmetric vibrations in lipid CH2 | Wallach et al., 1979; Casal and Mantsch, 1984 |
| 2850 | ν(C-H) symmetric vibrations in lipid CH2 | Wallach et al., 1979; Casal and Mantsch, 1984 |
| 1735 | ν(C=O) vibration from aliphatic esters | Wallach et al., 1979; Casal and Mantsch, 1984; Dubis et al., 1999 |
| 1712 | ν(C=O) vibration from aliphatic ketones | Wallach et al., 1979; Casal and Mantsch, 1984; Dubis et al., 1999 |
| 1471 and 1461 | δ(C-H) mode in lipid CH2 | Wallach et al., 1979; Casal and Mantsch, 1984; Merk et al., 1997 |
| 1446 | δ(C-H) mode adjacent to C=O | Spiker Jr et al., 1976; Wallach et al., 1979 |
| 1380 | ν(C-O) vibration from aromatic C-O , δ(C-H) symmetric in lipid CH3 | Wallach et al., 1979; Casal and Mantsch, 1984 |
| 1174 | δ(C-H) mode in CH2, ν(C-O) asymmetric vibration for esters | Spiker Jr et al., 1976; Wallach et al., 1979; Casal and Mantsch, 1984 |
| 977 | ν(C-N) asymmetric vibration in lipid N(CH3)3 | Wallach et al., 1979; Casal and Mantsch, 1984 |
| 729 and 719 | ρ(C-H) vibration in lipid CH2 | Wallach et al., 1979; Casal and Mantsch, 1984; Merk et al., 1997 |
Casal, H.L., Mantsch, H.H. (1984) Polymorphic phase behaviour of phospholipid membranes studied by infrared spectroscopy. Biochimica et Biophysica Acta (BBA) - Reviews on Biomembranes 779, 381-401. doi: 0.1016/0304-4157(84)90017-0.
Dubis, E.N., Dubis, A.T., Morzycki, J.W. (1999) Comparative analysis of plant cuticular waxes using HATR FT-IR reflection technique. Journal of Molecular Structure 511–512, 173-179. doi: 10.1016/S0022-2860(99)00157-X.
Merk, S., Blume, A., Riederer, M. (1997) Phase behaviour and crystallinity of plant cuticular waxes studied by Fourier transform infrared spectroscopy. Planta 204, 44-53. doi:10.1007/s004250050228.
Spiker Jr, R.C., T.J. Pinnavala and I.W. Levin. 1976. Use of phospholipid-clay complexes for determining vibrational spectra of membrane related systems. Biochimica et Biophysica Acta (BBA) - Biomembranes 455, 588-596. doi: 10.1016/0005-2736(76)90327-8.
Wallach, D.F.H., Verma, S.P., Fookson, J. (1979) Application of laser Raman and infrared spectroscopy to the analysis of membrane structure. Biochimica et Biophysica Acta (BBA) - Reviews on Biomembranes 559, 153-208. doi: 10.1016/0304-4157(79)90001-7.
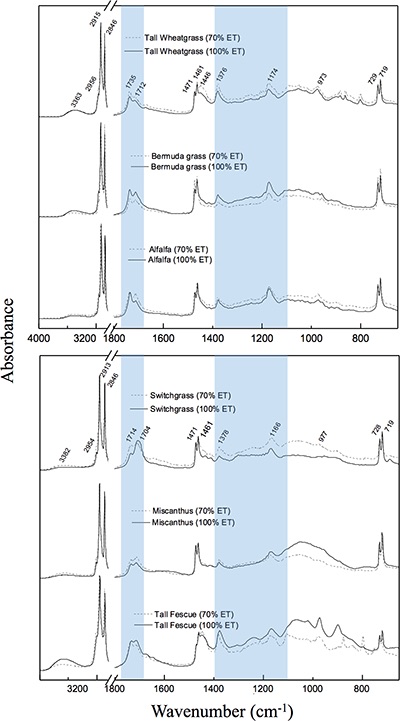
Figure S-1 Infrared spectra by species and treatment. Highlighted areas correspond to oxygen-containing groups depicted in Table S-3. Differences between full water replenished (solid lines) and deficit irrigation (dashed lines) were used to generate the drought response spectra shown in Figure 2.
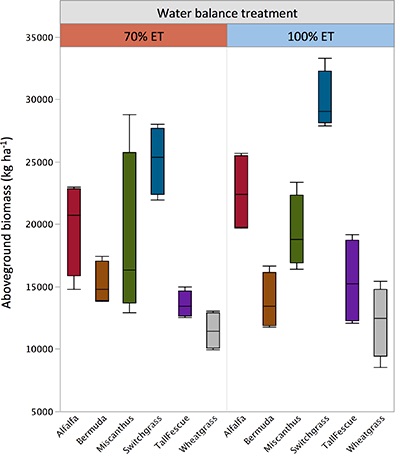
Figure S-2 Total aboveground biomass by species and treatment. Horizontal lines within the boxes represent median values. The ends of the boxes represent the 75th and 25th quantiles and whiskers span the entire data set including outliers.
Figures and Tables
Table 1 Species description.
| Species | Scientific name | Variety | Metabolim |
| Miscanthus | Miscanthus giganteus | Hybrid giganteus | Warm-season C4 |
| Switchgrass | Panicum virgatum | Alamo | Warm-season C4 |
| Bermudagrass | Cynodon dactylon | Giant NK 37 | Warm-season C4 |
| Alfalfa | Medicago sativa | Big Kaw | Warm-season C3 |
| Tall Fescue | Festuca arundinacea | Fawn | Cool-season C3 |
| Tall wheatgrass | Agropyron elongatum | Jose | Cool-season C3 |
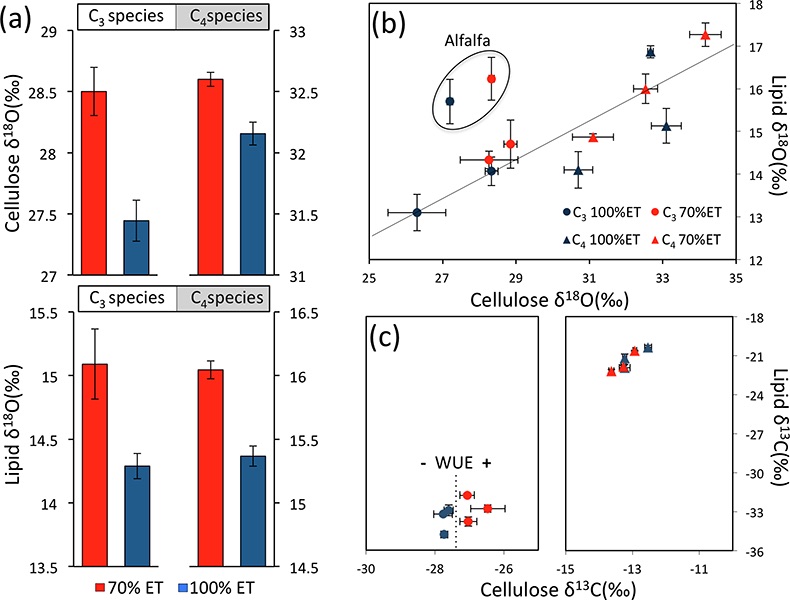
Figure 1 (a) Average δ18O values of C3 and C4 species under full evapotranspiration replenishment (100 % ET) and deficit irrigation (70 % ET). Differences between treatments were significant (t-test; P < 0.01) in all cases. (b) Relationship between cellulose and lipid δ18O values excluding alfalfa: y = 0.4449x + 1.4277; R² = 0.76 (P < 0.01). (c) Cellulose and lipid δ13C values; significant differences were observed in cellulose of C3 species (t-test; P < 0.01), but not in lipids. Error bars represent one standard deviation.
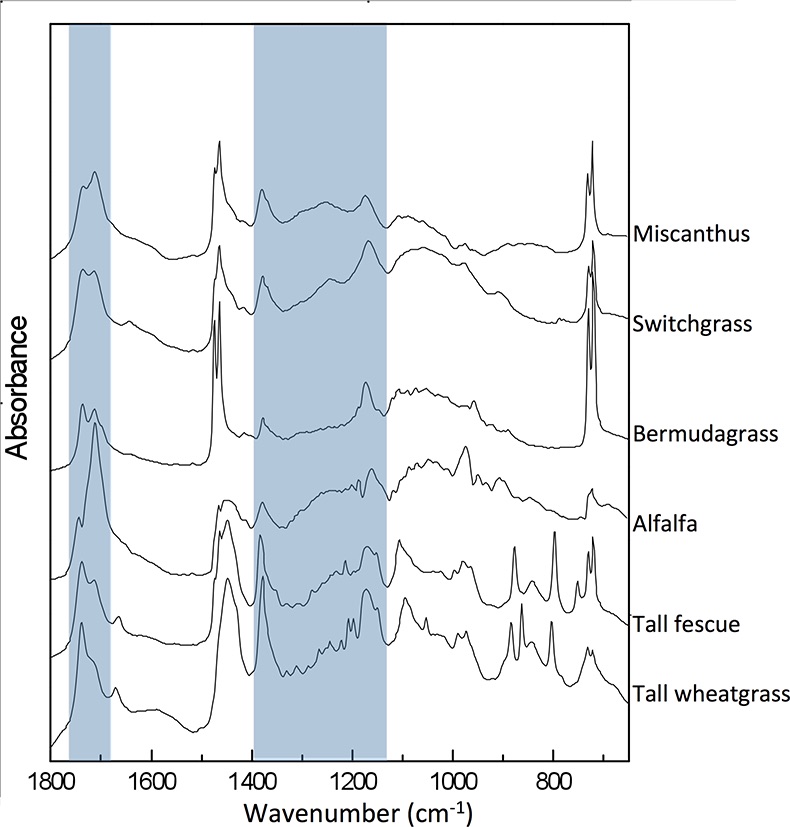
Figure 2 Difference spectra obtained by subtracting the spectra of lipid extracts of plants growing under deficit irrigation (70 % ET) from those growing under full water replenishment. Highlighted areas correspond to oxygen-containing groups (e.g., vibration from aromatic C-O, aliphatic esters and ketones). The spectra have been shifted vertically for clarity. A complete spectral characterisation is presented in Table S-3 and Figure S-1.
Back to article
Supplementary Figures and Tables
Table S-1 Climate at the study location.
| Climatic characteristics | Five Points |
| Köppen climate classification | Cool semi-arid climate |
| Average annual temp. (°C) | 17.1 |
| Average max. annual temp. (°C) | 25 |
| Average min. annual temp. (°C) | 9.2 |
| Average annual precipitation (mm) | 173 |
| Frost free days | 335 |
| Altitude (m) | 70 |
Table S-2 Soil properties at the study location.
| Soil Properties | |
| Clay (g kg-1) | 310 |
| Silt (g kg-1) | 340 |
| Sand (g kg-1) | 350 |
| pH (saturated paste extract) | 7.6 |
| CEC (cmolc kg-1) | 30.7 |
| Olsen-P (mg kg-1) | 7.4 |
| NO3-N (mg kg-1) | 10,6 |
| Extractable K (mg kg-1) | 439 |
| Organic matter (g kg-1) | 9.5 |
| Total N (g kg-1) | 0.7 |
Table S-3 Peaks associated with O-containing groups in the infrared spectra.
| Wavenumber (cm-1) | Assignment | Reference |
| 3360 | ν(O-H) vibrations in OH | Dubis et al., 1999 |
| 2950 | ν(C-H) asymmetric vibrations in lipid CH3 | Wallach et al., 1979; Casal and Mantsch, 1984 |
| 2915 | ν(C-H) asymmetric vibrations in lipid CH2 | Wallach et al., 1979; Casal and Mantsch, 1984 |
| 2850 | ν(C-H) symmetric vibrations in lipid CH2 | Wallach et al., 1979; Casal and Mantsch, 1984 |
| 1735 | ν(C=O) vibration from aliphatic esters | Wallach et al., 1979; Casal and Mantsch, 1984; Dubis et al., 1999 |
| 1712 | ν(C=O) vibration from aliphatic ketones | Wallach et al., 1979; Casal and Mantsch, 1984; Dubis et al., 1999 |
| 1471 and 1461 | δ(C-H) mode in lipid CH2 | Wallach et al., 1979; Casal and Mantsch, 1984; Merk et al., 1997 |
| 1446 | δ(C-H) mode adjacent to C=O | Spiker Jr et al., 1976; Wallach et al., 1979 |
| 1380 | ν(C-O) vibration from aromatic C-O , δ(C-H) symmetric in lipid CH3 | Wallach et al., 1979; Casal and Mantsch, 1984 |
| 1174 | δ(C-H) mode in CH2, ν(C-O) asymmetric vibration for esters | Spiker Jr et al., 1976; Wallach et al., 1979; Casal and Mantsch, 1984 |
| 977 | ν(C-N) asymmetric vibration in lipid N(CH3)3 | Wallach et al., 1979; Casal and Mantsch, 1984 |
| 729 and 719 | ρ(C-H) vibration in lipid CH2 | Wallach et al., 1979; Casal and Mantsch, 1984; Merk et al., 1997 |
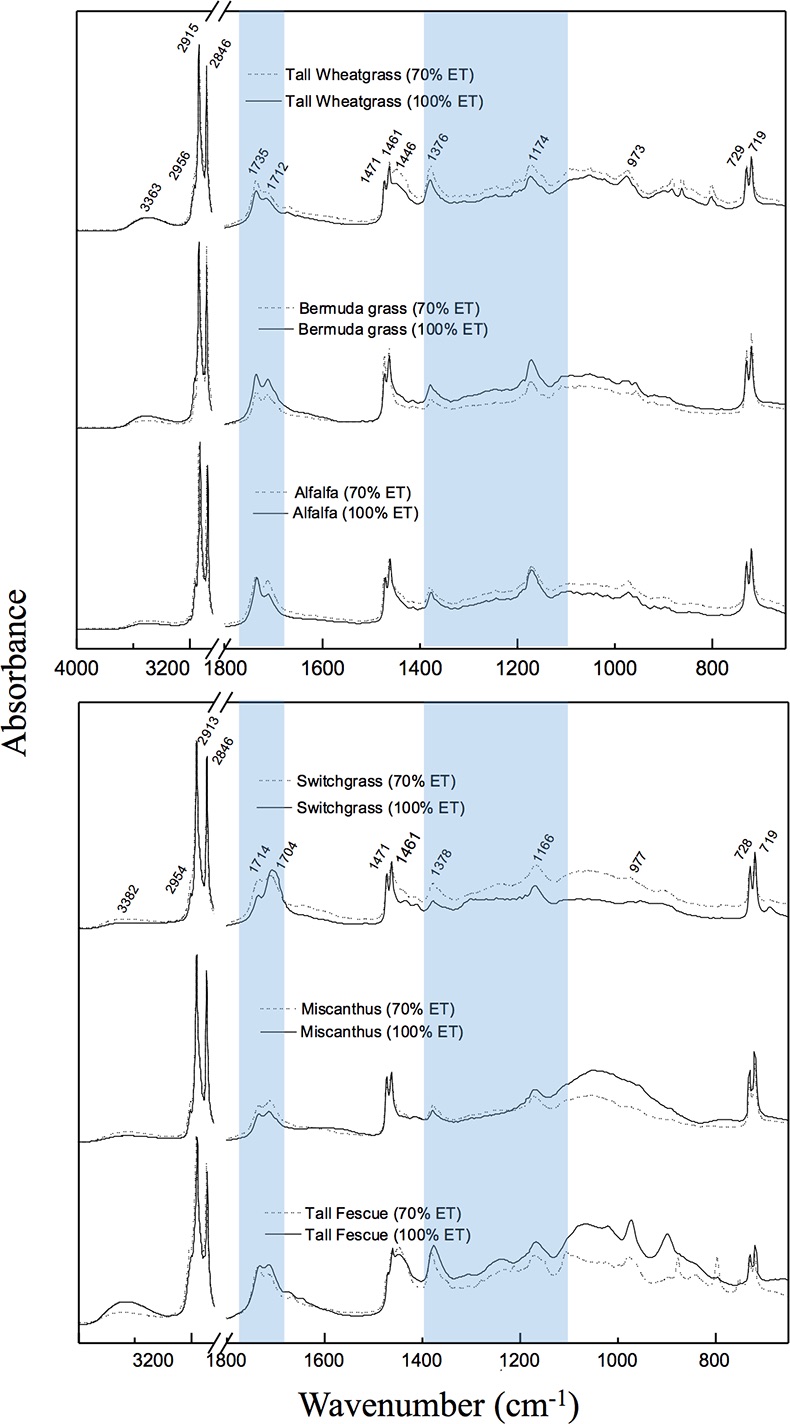
Figure S-1 Infrared spectra by species and treatment. Highlighted areas correspond to oxygen-containing groups depicted in Table S-3. Differences between full water replenished (solid lines) and deficit irrigation (dashed lines) were used to generate the drought response spectra shown in Figure 2.
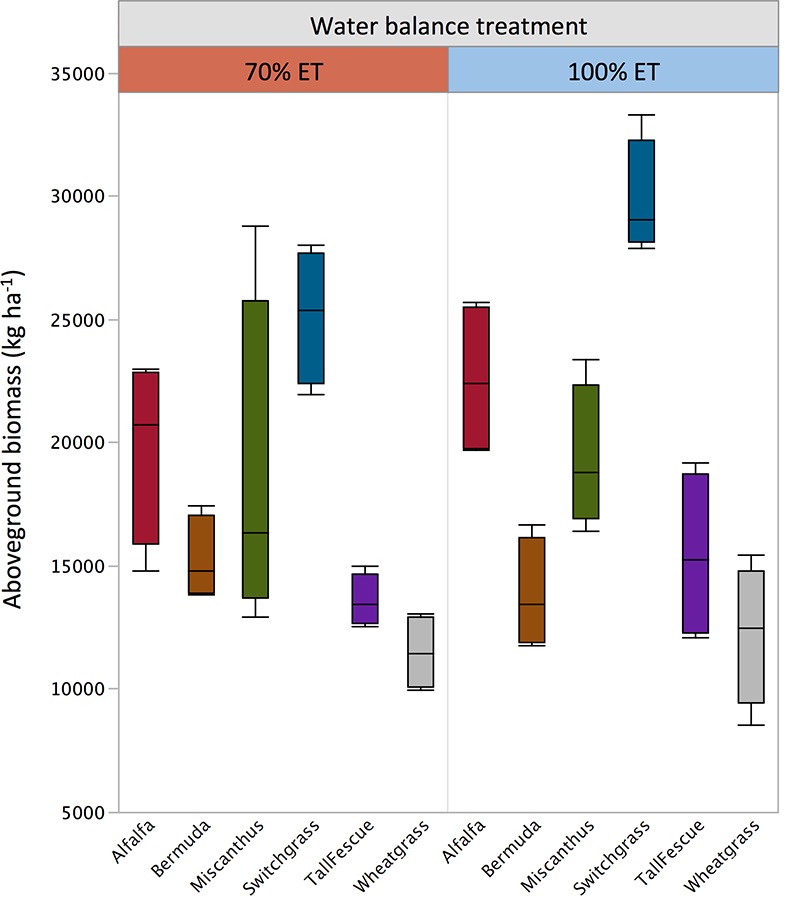
Figure S-2 Total aboveground biomass by species and treatment. Horizontal lines within the boxes represent median values. The ends of the boxes represent the 75th and 25th quantiles and whiskers span the entire data set including outliers.