



Nitrogen isotope signatures of microfossils suggest aerobic metabolism 3.0 Gyr ago
Affiliations | Corresponding Author | Cite as | Funding informationPublished by the European Association of Geochemistry
under Creative Commons License CC BY-NC-ND 4.0
Keywords: ammonia, early life, Farrel quartzite, microfossils, NanoSIMS

- Share this article





Article views:6,173Cumulative count of HTML views and PDF downloads.
- Download Citation
- Rights & Permissions

top
Abstract
Figures and Tables
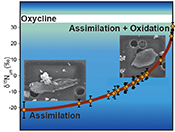
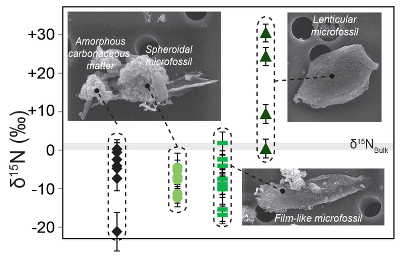 Figure 1 Nitrogen isotopic composition of amorphous carbonaceous matter, and of spheroidal, film-like and lenticular-like microfossils from the 3.0 Ga Farrel Quartzite formation. δ15NBulk values are provided for comparison with δ15Nµm values. | 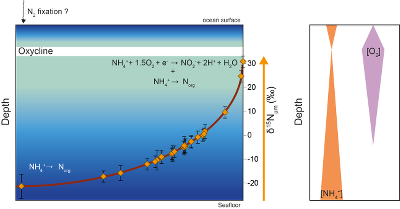 Figure 2 Cartoon illustrating the sedimentary conditions of the Farrel Quartzite formation. The blue gradient in the water column indicates change in NH4+ concentration from high (dark) to low (light) concentrations in anoxic zones. δ15Nµm values determined at the scale of individual microfossil and organic particle are represented by orange diamonds. In the proposed model, negative δ15Nµm values reflect assimilation of NH4+ from benthic efflux whereas positive δ15Nµm values reflect 15N Rayleigh distillation of NH4+ upward through ammonia oxidation when NH4+ concentrations were too low to allow the isotopic fractionation of ammonia assimilation to be expressed (see main text). |
| Figure 1 | Figure 2 |
top
Introduction
The isotopic composition of nitrogen (N) has been investigated in order to tackle questions related to Archean life (Beaumont and Robert, 1999
Beaumont, V., Robert, F. (1999) Nitrogen isotope ratios of kerogens in Precambrian cherts: a record of the evolution of atmosphere chemistry? Precambrian Research 96, 63-82.
; Pinti et al., 2001Pinti, D.L., Hashizume, K., Matsuda, J. (2001) Nitrogen and argon signatures in 3.8 to 2.8 Ga metasediments: Clues on the chemical state of the Archean ocean and the deep biosphere. Geochimica et Cosmochimica Acta 65, 2301-2315.
; Thomazo et al., 2011Thomazo, C., Ader, M., Philippot, P. (2011) Extreme 15N-enrichments in 2.72-Gyr-old sediments: evidence for a turning point in the nitrogen cycle. Geobiology 9, 107-120.
; Stueken et al., 2015aStueken, E.E., Buick, R., Guy, B.M., Koehler, M.C. (2015a) Isotopic evidence for biological nitrogen fixation by molybdenum-nitrogenase from 3.2 Gyr. Nature 520, 666-669.
). In the present day natural environment, N is mostly present as di-nitrogen (N2), ammonium (NH4+), organic N, nitrite (NO2-) and nitrate (NO3-). Within the marine N cycle, N compounds are closely coupled in a redox reaction network including specific isotope fractionation processes (Sigman et al., 2009Sigman, D.M., Karsh, K.L., Casciotti, K.L. (2009) Nitrogen isotopes in the ocean. In: Steele J.H., Thorpe, S.A., Turekian, K.K. (Eds.) Encyclopedia of Ocean Sciences. Academic Press, Oxford, 40-54.
). Accordingly, δ15N of sedimentary organic matter (OM) has been used to infer the evolution of the redox conditions of Archean oceans. For instance, negative δ15N values ranging between -7 and 0 ‰ in the OM isolated from early Archean rocks (3.6 to 3.2 Ga) have been interpreted as reflecting (i) anoxic Archean oceans in which microorganisms consumed N2 and NH4+ (Beaumont and Robert, 1999Beaumont, V., Robert, F. (1999) Nitrogen isotope ratios of kerogens in Precambrian cherts: a record of the evolution of atmosphere chemistry? Precambrian Research 96, 63-82.
) or (ii) a consumption of N2 and NH4+ by chemosynthetic bacteria flourishing close to 15N-depleted hydrothermal vents (Pinti et al., 2001Pinti, D.L., Hashizume, K., Matsuda, J. (2001) Nitrogen and argon signatures in 3.8 to 2.8 Ga metasediments: Clues on the chemical state of the Archean ocean and the deep biosphere. Geochimica et Cosmochimica Acta 65, 2301-2315.
; Shen et al., 2006Shen, Y., Pinti, D.L., Hashizume, K. (2006) Biogeochemical cycles of sulfur and nitrogen in the archean ocean and atmosphere. Archean Geodynamics and Environments 164, 305-320.
). The late Archean era (2.8 to 2.5 Ga) is characterised by the occurrence of positive OM δ15N values (up to ca. +5 ‰) presumably reflecting a rise in the abundance of oxidised N species (Beaumont and Robert, 1999Beaumont, V., Robert, F. (1999) Nitrogen isotope ratios of kerogens in Precambrian cherts: a record of the evolution of atmosphere chemistry? Precambrian Research 96, 63-82.
; Garvin et al., 2009Garvin, J., Buick, R., Anbar, A.D., Arnold, G.L., Kaufman, A.J. (2009) Isotopic Evidence for an Aerobic Nitrogen Cycle in the Latest Archean. Science 323, 1045-1048.
; Godfrey and Falkowski, 2009Godfrey, L.V., Falkowski, P.G. (2009) The cycling and redox state of nitrogen in the Archaean ocean. Nature Geoscience 2, 725-729.
). This suggests that significant free O2 was present in the oceans at that time, which is also supported by several inorganic palaeoredox proxies (Anbar et al., 2007Anbar, A.D., Duan, Y., Lyons, T.W., Arnold, G.L., Kendall, B., Creaser, R.A., Kaufman, A.J., Gordon, G.W., Scott, C., Garvin, J., Buick, R. (2007) A whiff of oxygen before the Great Oxidation Event? Science 317, 1903-1906.
; Reinhard et al., 2009Reinhard, C.T., Raiswell, R., Scott, C., Anbar, A.D., Lyons, T.W. (2009) A Late Archean Sulfidic Sea Stimulated by Early Oxidative Weathering of the Continents. Science 326, 713-716.
; Stueken et al., 2015bStueken, E.E., Buick, R., Anbar, A.D. (2015b) Selenium isotopes support free O-2 in the latest Archean. Geology 43, 259-262.
).Although early and late Archean sedimentary δ15N values suggest a progressive rise in the oxygenation level of Archean oceans, mid-Archean times (3.2 to 2.8 Ga) are characterised by a narrow range of δ15N values close to the N isotope composition of the present day atmosphere (0 ‰), suggesting the dominance of N2 fixing microorganisms (Stueken et al., 2015a
Stueken, E.E., Buick, R., Guy, B.M., Koehler, M.C. (2015a) Isotopic evidence for biological nitrogen fixation by molybdenum-nitrogenase from 3.2 Gyr. Nature 520, 666-669.
). However, complementary geochemical evidence points toward oxygenic photosynthesis occurring as early as ca. 3.0 Ga (Crowe et al., 2013Crowe, S.A., Dossing, L.N., Beukes, N.J., Bau, M., Kruger, S.J., Frei, R., Canfield, D.E. (2013) Atmospheric oxygenation three billion years ago. Nature 501, 535-538.
; Lyons et al., 2014Lyons, T.W., Reinhard, C.T., Planavsky, N.J. (2014) The rise of oxygen in Earth's early ocean and atmosphere. Nature 506, 307-315.
; Planavsky et al., 2014Planavsky, N.J., Asael, D., Hofmann, A., Reinhard, C.T., Lalonde, S.V., Knudsen, A., Wang, X.L., Ossa, F.O., Pecoits, E., Smith, A.J.B., Beukes, N.J., Bekker, A., Johnson, T.M., Konhauser, K.O., Lyons, T.W., Rouxel, O.J. (2014) Evidence for oxygenic photosynthesis half a billion years before the Great Oxidation Event. Nature Geoscience 7, 283-286.
) and ocean redox stratification ca. 3.2 Gyr ago (Satkoski et al., 2015Satkoski, A.M., Beukes, N.J., Li, W.Q., Beard, B.L., Johnson, C.M. (2015) A redox-stratified ocean 3.2 billion years ago. Earth and Planetary Science Letters 430, 43-53.
). If correct, such early oxygenation and ocean redox stratification should be recorded in redox-dependent sedimentary δ15N values, implying in turn that the δ15N values around 0 ± 2 ‰ measured on bulk samples might mask isotopic heterogeneities present at smaller scales. To investigate this issue, we have characterised the N isotope composition of organic microfossils (noted δ15Nµm) isolated from the mid-Archean Farrel Quartzite formation (3.0 Ga; Sugitani et al., 2007Sugitani, K., Grey, K., Allwood, A., Nagaoka, T., Mimura, K., Minami, M., Marshall, C.P., Van Kranendonk, M.J., Walter, M.R. (2007) Diverse microstructures from Archaean chert from the mount goldsworthy-mount grant area, pilbara craton, western australia: Microfossils, dubiofossils, or pseudofossils? Precambrian Research 158, 228-262.
) using nanoscale secondary ion mass spectrometry (NanoSIMS).top
Material and Methods
Organic-walled microfossils were isolated by acid maceration using a mixture of hydrofluoric and hydrochloric acids. In the acid maceration residue, we identified lenticular, spheroid- and film-like microfossils following the previously reported taxonomy (Sugitani et al., 2007
Sugitani, K., Grey, K., Allwood, A., Nagaoka, T., Mimura, K., Minami, M., Marshall, C.P., Van Kranendonk, M.J., Walter, M.R. (2007) Diverse microstructures from Archaean chert from the mount goldsworthy-mount grant area, pilbara craton, western australia: Microfossils, dubiofossils, or pseudofossils? Precambrian Research 158, 228-262.
; see also Supplementary Information). In the absence of unequivocal morphological features, the other organic particles were assigned to the “amorphous carbonaceous matter” group.From a morphological point of view, previous investigations indicate that acid maceration does not affect the morphological integrity of organic-walled microfossils from the Farrel Quartzite (Grey and Sugitani, 2009
Grey, K., Sugitani, K. (2009) Palynology of Archean microfossils (c. 3.0 Ga) from the Mount Grant area, Pilbara Craton, Western Australia: Further evidence of biogenicity. Precambrian Research 173, 60-69.
; Delarue et al., 2017Delarue, F., Robert, F., Sugitani, K., Tartese, R., Duhamel, R., Derenne, S. (2017) Investigation of the Geochemical Preservation of ca. 3.0 Ga Permineralized and Encapsulated Microfossils by Nanoscale Secondary Ion Mass Spectrometry. Astrobiology 17, 1192-1202.
). From a geochemical point of view, preservation of N content is highly variable across individual microfossils isolated from the Farrel Quartzite (Delarue et al., 2017Delarue, F., Robert, F., Sugitani, K., Tartese, R., Duhamel, R., Derenne, S. (2017) Investigation of the Geochemical Preservation of ca. 3.0 Ga Permineralized and Encapsulated Microfossils by Nanoscale Secondary Ion Mass Spectrometry. Astrobiology 17, 1192-1202.
). This suggests that acid maceration does not cause any significant hydrolysis of N-bearing compounds, which would likely have homogenised the N/C atomic ratios of microfossils through the preferential degradation of chemically labile N organic compounds. Similarly, large variations in δ15Nµm are observed among the microfossils, which can only be accounted for by Rayleigh distillation. The latter cannot have occurred during the acid treatment, as N isotope exchange through Rayleigh distillation cannot take place between solids (N in microfossils) and a fluid (N dissolved organic compounds in acid solution). Therefore, it is unlikely that acid maceration induced significant modification of both N/C atomic ratios and N isotopic compositions determined at the scale of individual microfossil by NanoSIMS.top
Results
The bulk nitrogen isotope composition (noted δ15NBulk) measured on Farrel Quartzite OM ranges between +0.3 ‰ and +2.2 ‰ (see Supplementary Information; n = 3; mean δ15NBulk = 1.0 ± 1.1 ‰; Fig. 1 and Table S-2). These δ15NBulk values are consistent with the average δ15N value calculated from all values determined by NanoSIMS on amorphous carbonaceous matter and microfossils isolated from the mineral matrix (δ15Nµm = -3.9 ± 11.5 ‰; n = 27; Fig. 1 and Table S-2). However, at the microscale, the δ15Nµm values measured on amorphous carbonaceous matter and microfossils vary from -21.6 ± 5.1 ‰ to +30.7 ± 2.3 ‰. Amorphous carbonaceous matter alone shows δ15Nµm values ranging between -21.6 ± 5.1 ‰ and 0.0 ± 2.5 ‰ (n = 8; Fig. 1). Film-like (n = 9) and spheroid-like microfossils (n = 6) exhibit mostly negative δ15Nµm values ranging from -17.2 ± 2.3 ‰ to +1.6 ± 2.9 ‰, while lenticular microfossils are characterised by positive δ15Nµm values ranging from +0.5 ± 2.4 ‰ to +30.7 ± 2.3 ‰ (n = 4; Fig. 1). Based on their nitrogen isotope composition, there is thus a clear discrimination between lenticular microfossils on the one hand and spheroid- and film-like microfossils on the other hand.
Figure 1 Nitrogen isotopic composition of amorphous carbonaceous matter, and of spheroidal, film-like and lenticular-like microfossils from the 3.0 Ga Farrel Quartzite formation. δ15NBulk values are provided for comparison with δ15Nµm values.
top
Discussion
The quasi-null δ15NBulk values measured on Farrel Quartzite OM hide unexpectedly large δ15Nµm variations in isolated microfossils (Fig. 1 and Table S-2). The average of δ15Nµm values is close to δ15NBulk. This observation can be explained if the bulk value integrates a diversity of organic particles with variable δ15N.
Before relating measured δ15Nµm values to potential metabolic effects, it is a prerequisite to ensure that biostratonomic and fossilisation processes did not significantly alter the original N isotopic composition. Biomass degradation can cause δ15N variations of ca. 3-4 ‰ (Lehmann et al., 2002
Lehmann, M.F., Bernasconi, S.M., Barbieri, A., McKenzie, J.A. (2002) Preservation of organic matter and alteration of its carbon and nitrogen isotope composition during simulated and in situ early sedimentary diagenesis. Geochimica et Cosmochimica Acta 66, 3573-3584.
), while metamorphism up to greenschist facies, as underwent by the Farrel Quartzite samples, can only induce restricted δ15N modification of up to ~1 to 2 ‰ (Ader et al., 2006Ader, M., Cartigny, P., Boudou, J.P., Oh, J.H., Petit, E., Javoy, M. (2006) Nitrogen isotopic evolution of carbonaceous matter during metamorphism: Methodology and preliminary results. Chemical Geology 232, 152-169.
; see Supplementary Information for further details). It thus seems unlikely that secondary processes have largely modified δ15Nµm values, which, in turn, may be linked with N metabolism(s) in the water column.In the N cycle, only Rayleigh fractionation has been reported to yield large variations of δ15N values through the partial consumption of NO3- or of NH4+, leaving the residual dissolved inorganic nitrogen (DIN) enriched in 15N (Sigman et al., 2009
Sigman, D.M., Karsh, K.L., Casciotti, K.L. (2009) Nitrogen isotopes in the ocean. In: Steele J.H., Thorpe, S.A., Turekian, K.K. (Eds.) Encyclopedia of Ocean Sciences. Academic Press, Oxford, 40-54.
). As the NO3- concentration was negligible in Archean oceans (Falkowski and Godfrey, 2008Falkowski, P.G., Godfrey, L.V. (2008) Electrons, life and the evolution of Earth's oxygen cycle. Philosophical Transactions of the Royal Society B-Biological Sciences 363, 2705-2716.
), it is unlikely that partial conversion of NO3- into N2 occurred and gave rise to the observed δ15Nµm variations. Thus, only the partial consumption of NH4+ seems to be an acceptable mechanism to explain the large variation in δ15Nµm values, consistent with an Archean DIN marine reservoir dominated by NH4+ (Thomazo et al., 2011Thomazo, C., Ader, M., Philippot, P. (2011) Extreme 15N-enrichments in 2.72-Gyr-old sediments: evidence for a turning point in the nitrogen cycle. Geobiology 9, 107-120.
). Three different marine metabolisms (referred to as chemolithoautotrophy) could then be involved in the nitrogen isotope cycle: (i) NH4+ assimilation (NH4+ → Norg), (ii) anaerobic NH4+ oxidation (anammox; NH4+ + NO2- → N2 + 2H2O) and (iii) aerobic NH4+ oxidation (NH4+ + 1.5 O2 → NO2- + 2H+ + H2O). The N isotope fractionations (noted ε, with ε ≈ δ15NNH4+ - δ15NProduct) associated with these processes have been determined experimentally, and range from +4 ‰ ≤ ε ≤ +27 ‰ for (i) and +14 ‰ ≤ ε ≤ +19 ‰ for (iii), respectively (Hoch et al., 1992Hoch, M.P., Fogel, M.L., Kirchman, D.L. (1992) Isotope fractionation associated with ammonium uptake by a marine bacterium. Limnology and Oceanography 37, 1447-1459.
; Casciotti et al., 2003Casciotti, K.L., Sigman, D.M., Ward, B.B. (2003) Linking diversity and stable isotope fractionation in ammonia-oxidizing bacteria. Geomicrobiology Journal 20, 335-353.
). Anammox is not further considered here since its antiquity is still a controversial issue and the resulting δ15NBulk remains unknown (Stueken et al., 2016Stueken, E.E., Kipp, M.A., Koehler, M.C., Buick, R. (2016) The evolution of Earth's biogeochemical nitrogen cycle. Earth-Science Reviews 160, 220-239.
; see also Supplementary Information).In modern environmental settings, δ15N variations rarely show large variations. However, in the modern Black Sea environment, large δ15N variations ranging from negative up to positive values have been observed to be associated with chemolithoautotrophy along a spatial redox gradient that promotes the progressive oxidation of NH4+ in the deep and suboxic level of the Black Sea (Coban-Yildiz et al., 2006
Coban-Yildiz, Y., Altabet, M.A., Yilmaz, A., Tugrul, S. (2006) Carbon and nitrogen isotopic ratios of suspended particulate organic matter (SPOM) in the Black Sea water column. Deep-Sea Research Part II-Topical Studies in Oceanography 53, 1875-1892.
). According to this observation, we propose that mineralisation of OM yielded large amounts of NH4+ in the deeper level of the ocean (Fig. 2). Preferential assimilation of 14NH4+ from this benthic efflux may explain the negative δ15Nµm values measured on spheroids and film/filament-like microfossils. The latter are thus likely remnants of benthic microbial mats, in agreement with their morphology (Westall et al., 2006Westall, F., de Ronde, C.E.J., Southam, G., Grassineau, N., Colas, M., Cockell, C.S., Lammer, H. (2006) Implications of a 3.472-3.333 Gyr-old subaerial microbial mat from the Barberton greenstone belt, South Africa for the UV environmental conditions on the early Earth. Philosophical Transactions of the Royal Society B-Biological Sciences 361, 1857-1875.
).Figure 2 Cartoon illustrating the sedimentary conditions of the Farrel Quartzite formation. The blue gradient in the water column indicates change in NH4+ concentration from high (dark) to low (light) concentrations in anoxic zones. δ15Nµm values determined at the scale of individual microfossil and organic particle are represented by orange diamonds. In the proposed model, negative δ15Nµm values reflect assimilation of NH4+ from benthic efflux whereas positive δ15Nµm values reflect 15N Rayleigh distillation of NH4+ upward through ammonia oxidation when NH4+ concentrations were too low to allow the isotopic fractionation of ammonia assimilation to be expressed (see main text).
We propose that assimilation of NH4+ from benthic efflux in the deep water column led to both a decrease in NH4+ concentration and a progressive enrichment in 15NH4+ in the upper levels of the water column (Fig. 2). Partial NH4+ assimilation can yield ε values down to -27 ‰ when NH4+ concentration is higher than 20 μM, while lower NH4+ concentrations are associated with lower ε values close to -4 ‰ (see Stueken et al., 2016
Stueken, E.E., Kipp, M.A., Koehler, M.C., Buick, R. (2016) The evolution of Earth's biogeochemical nitrogen cycle. Earth-Science Reviews 160, 220-239.
and references therein). In this context, such lower concentrations are unlikely to yield δ15Nµm values up to +31 ‰ as recorded in the lenticular microfossils. These microfossils are interpreted as remnants of pelagic microorganisms based on both their morphology (Sugitani et al., 2007Sugitani, K., Grey, K., Allwood, A., Nagaoka, T., Mimura, K., Minami, M., Marshall, C.P., Van Kranendonk, M.J., Walter, M.R. (2007) Diverse microstructures from Archaean chert from the mount goldsworthy-mount grant area, pilbara craton, western australia: Microfossils, dubiofossils, or pseudofossils? Precambrian Research 158, 228-262.
; Oehler et al., 2010Oehler, D.Z., Robert, F., Walter, M.R., Sugitani, K., Meibom, A., Mostefaoui, S., Gibson, E.K. (2010) Diversity in the Archean Biosphere: New Insights from NanoSIMS. Astrobiology 10, 413-424.
) and their carbon isotope composition (House et al., 2013House, C.H., Oehler, D.Z., Sugitani, K., Mimura, K. (2013) Carbon isotopic analyses of ca. 3.0 Ga microstructures imply planktonic autotrophs inhabited Earth's early oceans. Geology 41, 651-654.
). We then propose that aerobic NH4+ oxidation drove 15N enrichment of NH4+ upward in the water column (Fig. 2). In this scenario, the positive δ15Nµm values might be regarded as a redox signal as previously reported (Coban-Yildiz et al., 2006Coban-Yildiz, Y., Altabet, M.A., Yilmaz, A., Tugrul, S. (2006) Carbon and nitrogen isotopic ratios of suspended particulate organic matter (SPOM) in the Black Sea water column. Deep-Sea Research Part II-Topical Studies in Oceanography 53, 1875-1892.
; Granger et al., 2011Granger, J., Prokopenko, M.G., Sigman, D.M., Mordy, C.W., Morse, Z.M., Morales, L.V., Sambrotto, R.N., Plessen, B. (2011) Coupled nitrification-denitrification in sediment of the eastern Bering Sea shelf leads to N-15 enrichment of fixed N in shelf waters. Journal of Geophysical Research-Oceans 116.
; Morales et al., 2014Morales, L.V., Granger, J., Chang, B.X., Prokopenko, M.G., Plessen, B., Gradinger, R., Sigman, D.M. (2014) Elevated N-15/N-14 in particulate organic matter, zooplankton, and diatom frustule-bound nitrogen in the ice-covered water column of the Bering Sea eastern shelf. Deep-Sea Research Part II-Topical Studies in Oceanography 109, 100-111.
). However, to be recorded in OM, aerobic NH4+ oxidation should have been coupled with NH4+ assimilation, leaving lenticular organic-walled microfossils enriched in 15N (Fig. 2).In view of the δ13C (House et al., 2013
House, C.H., Oehler, D.Z., Sugitani, K., Mimura, K. (2013) Carbon isotopic analyses of ca. 3.0 Ga microstructures imply planktonic autotrophs inhabited Earth's early oceans. Geology 41, 651-654.
) and of the present δ15N values along with morphological evidence, this proposed scenario, which combines morphological evidence with C and N isotopic data, suggests that life thrived across the entire water column down to the seafloor, and points to the early existence of aerobic metabolism 3 billion years ago. Since aerobic NH4+ oxidation implies the occurrence of free dissolved molecular O2 in the water column (Wang and Yang, 2004Wang, J.L., Yang, N. (2004) Partial nitrification under limited dissolved oxygen conditions. Process Biochemistry 39, 1223-1229.
), this finding echoes previous geochemical arguments pointing toward mid-Archean oxygen oases and, therefore, biological O2 production by oxygenic photosynthesis in stratified mid-Archean oceans (Crowe et al., 2013Crowe, S.A., Dossing, L.N., Beukes, N.J., Bau, M., Kruger, S.J., Frei, R., Canfield, D.E. (2013) Atmospheric oxygenation three billion years ago. Nature 501, 535-538.
; Lyons et al., 2014Lyons, T.W., Reinhard, C.T., Planavsky, N.J. (2014) The rise of oxygen in Earth's early ocean and atmosphere. Nature 506, 307-315.
; Planavsky et al., 2014Planavsky, N.J., Asael, D., Hofmann, A., Reinhard, C.T., Lalonde, S.V., Knudsen, A., Wang, X.L., Ossa, F.O., Pecoits, E., Smith, A.J.B., Beukes, N.J., Bekker, A., Johnson, T.M., Konhauser, K.O., Lyons, T.W., Rouxel, O.J. (2014) Evidence for oxygenic photosynthesis half a billion years before the Great Oxidation Event. Nature Geoscience 7, 283-286.
; Satkoski et al., 2015Satkoski, A.M., Beukes, N.J., Li, W.Q., Beard, B.L., Johnson, C.M. (2015) A redox-stratified ocean 3.2 billion years ago. Earth and Planetary Science Letters 430, 43-53.
).top
Acknowledgements
This research is supported by the ERC Grant No. 290861 – PaleoNanoLife (PI F. Robert). The National NanoSIMS Facility at the MNHN is supported by MNHN, CNRS, Region Ile de France, and Ministère de l’Enseignement supérieur et de la Recherche. Authors are grateful to Marie Balasse and Denis Fiorillo for bulk nitrogen isotopic composition analyses. We also thank three anonymous reviewers, Eva Stueken, Liane Benning and Ariel Anbar for their insightful comments.
Editor: Ariel Anbar
top
References
Ader, M., Cartigny, P., Boudou, J.P., Oh, J.H., Petit, E., Javoy, M. (2006) Nitrogen isotopic evolution of carbonaceous matter during metamorphism: Methodology and preliminary results. Chemical Geology 232, 152-169.
 Show in context
Show in context Biomass degradation can cause δ15N variations of ca. 3-4 ‰ (Lehmann et al., 2002), while metamorphism up to greenschist facies, as underwent by the Farrel Quartzite samples, can only induce restricted δ15N modification of up to ~1 to 2 ‰ (Ader et al., 2006; see Supplementary Information for further details).
View in article
Anbar, A.D., Duan, Y., Lyons, T.W., Arnold, G.L., Kendall, B., Creaser, R.A., Kaufman, A.J., Gordon, G.W., Scott, C., Garvin, J., Buick, R. (2007) A whiff of oxygen before the Great Oxidation Event? Science 317, 1903-1906.
Show in context This suggests that significant free O2 was present in the oceans at that time, which is also supported by several inorganic palaeoredox proxies (Anbar et al., 2007; Reinhard et al., 2009; Stueken et al., 2015b).
View in article
Beaumont, V., Robert, F. (1999) Nitrogen isotope ratios of kerogens in Precambrian cherts: a record of the evolution of atmosphere chemistry? Precambrian Research 96, 63-82.
Show in context The isotopic composition of nitrogen (N) has been investigated in order to tackle questions related to Archean life (Beaumont and Robert, 1999; Pinti et al., 2001; Thomazo et al., 2011; Stueken et al., 2015a).
View in article
For instance, negative δ15N values ranging between -7 and 0 ‰ in the OM isolated from early Archean rocks (3.6 to 3.2 Ga) have been interpreted as reflecting (i) anoxic Archean oceans in which microorganisms consumed N2 and NH4+ (Beaumont and Robert, 1999) or (ii) a consumption of N2 and NH4+ by chemosynthetic bacteria flourishing close to 15N-depleted hydrothermal vents (Pinti et al., 2001; Shen et al., 2006).
View in article
The late Archean era (2.8 to 2.5 Ga) is characterised by the occurrence of positive OM δ15N values (up to ca. +5 ‰) presumably reflecting a rise in the abundance of oxidised N species (Beaumont and Robert, 1999; Garvin et al., 2009; Godfrey and Falkowski, 2009).
View in article
Casciotti, K.L., Sigman, D.M., Ward, B.B. (2003) Linking diversity and stable isotope fractionation in ammonia-oxidizing bacteria. Geomicrobiology Journal 20, 335-353.
Show in context The N isotope fractionations (noted ε, with ε ≈ δ15NNH4+ - δ15NProduct) associated with these processes have been determined experimentally, and range from +4 ‰ ≤ ε ≤ +27 ‰ for (i) and +14 ‰ ≤ ε ≤ +19 ‰ for (iii), respectively (Hoch et al., 1992; Casciotti et al., 2003).
View in article
Coban-Yildiz, Y., Altabet, M.A., Yilmaz, A., Tugrul, S. (2006) Carbon and nitrogen isotopic ratios of suspended particulate organic matter (SPOM) in the Black Sea water column. Deep-Sea Research Part II-Topical Studies in Oceanography 53, 1875-1892.
Show in context However, in the modern Black Sea environment, large δ15N variations ranging from negative up to positive values have been observed to be associated with chemolithoautotrophy along a spatial redox gradient that promotes the progressive oxidation of NH4+ in the deep and suboxic level of the Black Sea (Coban-Yildiz et al., 2006).
View in article
In this scenario, the positive δ15Nµm values might be regarded as a redox signal as previously reported (Coban-Yildiz et al., 2006; Granger et al., 2011; Morales et al., 2014).
View in article
Crowe, S.A., Dossing, L.N., Beukes, N.J., Bau, M., Kruger, S.J., Frei, R., Canfield, D.E. (2013) Atmospheric oxygenation three billion years ago. Nature 501, 535-538.
Show in context However, complementary geochemical evidence points toward oxygenic photosynthesis occurring as early as ca. 3.0 Ga (Crowe et al., 2013; Lyons et al., 2014; Planavsky et al., 2014) and ocean redox stratification ca. 3.2 Gyr ago (Satkoski et al., 2015).
View in article
Since aerobic NH4+ oxidation implies the occurrence of free dissolved molecular O2 in the water column (Wang and Yang, 2004), this finding echoes previous geochemical arguments pointing toward mid-Archean oxygen oases and, therefore, biological O2 production by oxygenic photosynthesis in stratified mid-Archean oceans (Crowe et al., 2013; Lyons et al., 2014; Planavsky et al., 2014; Satkoski et al., 2015).
View in article
Delarue, F., Robert, F., Sugitani, K., Tartese, R., Duhamel, R., Derenne, S. (2017) Investigation of the Geochemical Preservation of ca. 3.0 Ga Permineralized and Encapsulated Microfossils by Nanoscale Secondary Ion Mass Spectrometry. Astrobiology 17, 1192-1202.
Show in context From a morphological point of view, previous investigations indicate that acid maceration does not affect the morphological integrity of organic-walled microfossils from the Farrel Quartzite (Grey and Sugitani, 2009; Delarue et al., 2017).
View in article
From a geochemical point of view, preservation of N content is highly variable across individual microfossils isolated from the Farrel Quartzite (Delarue et al., 2017).
View in article
Falkowski, P.G., Godfrey, L.V. (2008) Electrons, life and the evolution of Earth's oxygen cycle. Philosophical Transactions of the Royal Society B-Biological Sciences 363, 2705-2716.
Show in context As the NO3- concentration was negligible in Archean oceans (Falkowski and Godfrey, 2008), it is unlikely that partial conversion of NO3- into N2 occurred and gave rise to the observed δ15Nµm variations.
View in article
Garvin, J., Buick, R., Anbar, A.D., Arnold, G.L., Kaufman, A.J. (2009) Isotopic Evidence for an Aerobic Nitrogen Cycle in the Latest Archean. Science 323, 1045-1048.
Show in context The late Archean era (2.8 to 2.5 Ga) is characterised by the occurrence of positive OM δ15N values (up to ca. +5 ‰) presumably reflecting a rise in the abundance of oxidised N species (Beaumont and Robert, 1999; Garvin et al., 2009; Godfrey and Falkowski, 2009).
View in article
Godfrey, L.V., Falkowski, P.G. (2009) The cycling and redox state of nitrogen in the Archaean ocean. Nature Geoscience 2, 725-729.
Show in context The late Archean era (2.8 to 2.5 Ga) is characterised by the occurrence of positive OM δ15N values (up to ca. +5 ‰) presumably reflecting a rise in the abundance of oxidised N species (Beaumont and Robert, 1999; Garvin et al., 2009; Godfrey and Falkowski, 2009).
View in article
Granger, J., Prokopenko, M.G., Sigman, D.M., Mordy, C.W., Morse, Z.M., Morales, L.V., Sambrotto, R.N., Plessen, B. (2011) Coupled nitrification-denitrification in sediment of the eastern Bering Sea shelf leads to N-15 enrichment of fixed N in shelf waters. Journal of Geophysical Research-Oceans 116.
Show in context In this scenario, the positive δ15Nµm values might be regarded as a redox signal as previously reported (Coban-Yildiz et al., 2006; Granger et al., 2011; Morales et al., 2014).
View in article
Grey, K., Sugitani, K. (2009) Palynology of Archean microfossils (c. 3.0 Ga) from the Mount Grant area, Pilbara Craton, Western Australia: Further evidence of biogenicity. Precambrian Research 173, 60-69.
Show in context From a morphological point of view, previous investigations indicate that acid maceration does not affect the morphological integrity of organic-walled microfossils from the Farrel Quartzite (Grey and Sugitani, 2009; Delarue et al., 2017).
View in article
Hoch, M.P., Fogel, M.L., Kirchman, D.L. (1992) Isotope fractionation associated with ammonium uptake by a marine bacterium. Limnology and Oceanography 37, 1447-1459.
Show in context The N isotope fractionations (noted ε, with ε ≈ δ15NNH4+ - δ15NProduct) associated with these processes have been determined experimentally, and range from +4 ‰ ≤ ε ≤ +27 ‰ for (i) and +14 ‰ ≤ ε ≤ +19 ‰ for (iii), respectively (Hoch et al., 1992; Casciotti et al., 2003).
View in article
House, C.H., Oehler, D.Z., Sugitani, K., Mimura, K. (2013) Carbon isotopic analyses of ca. 3.0 Ga microstructures imply planktonic autotrophs inhabited Earth's early oceans. Geology 41, 651-654.
Show in context These microfossils are interpreted as remnants of pelagic microorganisms based on both their morphology (Sugitani et al., 2007; Oehler et al., 2010) and their carbon isotope composition (House et al., 2013).
View in article
In view of the δ13C (House et al., 2013) and of the present δ15N values along with morphological evidence, this proposed scenario, which combines morphological evidence with C and N isotopic data, suggests that life thrived across the entire water column down to the seafloor, and points to the early existence of aerobic metabolism 3 billion years ago.
View in article
Lehmann, M.F., Bernasconi, S.M., Barbieri, A., McKenzie, J.A. (2002) Preservation of organic matter and alteration of its carbon and nitrogen isotope composition during simulated and in situ early sedimentary diagenesis. Geochimica et Cosmochimica Acta 66, 3573-3584.
Show in context Biomass degradation can cause d15N variations of ca. 3-4 ‰ (Lehmann et al., 2002), while metamorphism up to greenschist facies, as underwent by the Farrel Quartzite samples, can only induce restricted δ15N modification of up to ~1 to 2 ‰ (Ader et al., 2006; see Supplementary Information for further details).
View in article
Lyons, T.W., Reinhard, C.T., Planavsky, N.J. (2014) The rise of oxygen in Earth's early ocean and atmosphere. Nature 506, 307-315.
Show in context However, complementary geochemical evidence points toward oxygenic photosynthesis occurring as early as ca. 3.0 Ga (Crowe et al., 2013; Lyons et al., 2014; Planavsky et al., 2014) and ocean redox stratification ca. 3.2 Gyr ago (Satkoski et al., 2015).
View in article
Since aerobic NH4+ oxidation implies the occurrence of free dissolved molecular O2 in the water column (Wang and Yang, 2004), this finding echoes previous geochemical arguments pointing toward mid-Archean oxygen oases and, therefore, biological O2 production by oxygenic photosynthesis in stratified mid-Archean oceans (Crowe et al., 2013; Lyons et al., 2014; Planavsky et al., 2014; Satkoski et al., 2015).
View in article
Morales, L.V., Granger, J., Chang, B.X., Prokopenko, M.G., Plessen, B., Gradinger, R., Sigman, D.M. (2014) Elevated N-15/N-14 in particulate organic matter, zooplankton, and diatom frustule-bound nitrogen in the ice-covered water column of the Bering Sea eastern shelf. Deep-Sea Research Part II-Topical Studies in Oceanography 109, 100-111.
Show in context In this scenario, the positive δ15Nµm values might be regarded as a redox signal as previously reported (Coban-Yildiz et al., 2006; Granger et al., 2011; Morales et al., 2014).
View in article
Oehler, D.Z., Robert, F., Walter, M.R., Sugitani, K., Meibom, A., Mostefaoui, S., Gibson, E.K. (2010) Diversity in the Archean Biosphere: New Insights from NanoSIMS. Astrobiology 10, 413-424.
Show in context These microfossils are interpreted as remnants of pelagic microorganisms based on both their morphology (Sugitani et al., 2007; Oehler et al., 2010) and their carbon isotope composition (House et al., 2013).
View in article
Pinti, D.L., Hashizume, K., Matsuda, J. (2001) Nitrogen and argon signatures in 3.8 to 2.8 Ga metasediments: Clues on the chemical state of the Archean ocean and the deep biosphere. Geochimica et Cosmochimica Acta 65, 2301-2315.
Show in context The isotopic composition of nitrogen (N) has been investigated in order to tackle questions related to Archean life (Beaumont and Robert, 1999; Pinti et al., 2001; Thomazo et al., 2011; Stueken et al., 2015a).
View in article
For instance, negative δ15N values ranging between -7 and 0 ‰ in the OM isolated from early Archean rocks (3.6 to 3.2 Ga) have been interpreted as reflecting (i) anoxic Archean oceans in which microorganisms consumed N2 and NH4+ (Beaumont and Robert, 1999) or (ii) a consumption of N2 and NH4+ by chemosynthetic bacteria flourishing close to 15N-depleted hydrothermal vents (Pinti et al., 2001; Shen et al., 2006).
View in article
Planavsky, N.J., Asael, D., Hofmann, A., Reinhard, C.T., Lalonde, S.V., Knudsen, A., Wang, X.L., Ossa, F.O., Pecoits, E., Smith, A.J.B., Beukes, N.J., Bekker, A., Johnson, T.M., Konhauser, K.O., Lyons, T.W., Rouxel, O.J. (2014) Evidence for oxygenic photosynthesis half a billion years before the Great Oxidation Event. Nature Geoscience 7, 283-286.
Show in context Reinhard, C.T., Raiswell, R., Scott, C., Anbar, A.D., Lyons, T.W. (2009) A Late Archean Sulfidic Sea Stimulated by Early Oxidative Weathering of the Continents. Science 326, 713-716.
Show in context This suggests that significant free O2 was present in the oceans at that time, which is also supported by several inorganic palaeoredox proxies (Anbar et al., 2007; Reinhard et al., 2009; Stueken et al., 2015b).
View in article
Satkoski, A.M., Beukes, N.J., Li, W.Q., Beard, B.L., Johnson, C.M. (2015) A redox-stratified ocean 3.2 billion years ago. Earth and Planetary Science Letters 430, 43-53.
Show in context However, complementary geochemical evidence points toward oxygenic photosynthesis occurring as early as ca. 3.0 Ga (Crowe et al., 2013; Lyons et al., 2014; Planavsky et al., 2014) and ocean redox stratification ca. 3.2 Gyr ago (Satkoski et al., 2015).
View in article
Since aerobic NH4+ oxidation implies the occurrence of free dissolved molecular O2 in the water column (Wang and Yang, 2004), this finding echoes previous geochemical arguments pointing toward mid-Archean oxygen oases and, therefore, biological O2 production by oxygenic photosynthesis in stratified mid-Archean oceans (Crowe et al., 2013; Lyons et al., 2014; Planavsky et al., 2014; Satkoski et al., 2015).
View in article
Shen, Y., Pinti, D.L., Hashizume, K. (2006) Biogeochemical cycles of sulfur and nitrogen in the archean ocean and atmosphere. Archean Geodynamics and Environments 164, 305-320.
Show in context For instance, negative δ15N values ranging between -7 and 0 ‰ in the OM isolated from early Archean rocks (3.6 to 3.2 Ga) have been interpreted as reflecting (i) anoxic Archean oceans in which microorganisms consumed N2 and NH4+ (Beaumont and Robert, 1999) or (ii) a consumption of N2 and NH4+ by chemosynthetic bacteria flourishing close to 15N-depleted hydrothermal vents (Pinti et al., 2001; Shen et al., 2006).
View in article
Sigman, D.M., Karsh, K.L., Casciotti, K.L. (2009) Nitrogen isotopes in the ocean. In: Steele J.H., Thorpe, S.A., Turekian, K.K. (Eds.) Encyclopedia of Ocean Sciences. Academic Press, Oxford, 40-54.
Show in context Within the marine N cycle, N compounds are closely coupled in a redox reaction network including specific isotope fractionation processes (Sigman et al., 2009).
View in article
In the N cycle, only Rayleigh fractionation has been reported to yield large variations of δ15N values through the partial consumption of NO3- or of NH4+, leaving the residual dissolved inorganic nitrogen (DIN) enriched in 15N (Sigman et al., 2009).
View in article
Stueken, E.E., Buick, R., Guy, B.M., Koehler, M.C. (2015a) Isotopic evidence for biological nitrogen fixation by molybdenum-nitrogenase from 3.2 Gyr. Nature 520, 666-669.
Show in context The isotopic composition of nitrogen (N) has been investigated in order to tackle questions related to Archean life (Beaumont and Robert, 1999; Pinti et al., 2001; Thomazo et al., 2011; Stueken et al., 2015a).
View in article
Although early and late Archean sedimentary δ15N values suggest a progressive rise in the oxygenation level of Archean oceans, mid-Archean times (3.2 to 2.8 Ga) are characterised by a narrow range of δ15N values close to the N isotope composition of the present day atmosphere (0 ‰), suggesting the dominance of N2 fixing microorganisms (Stueken et al., 2015a).
View in article
Stueken, E.E., Buick, R., Anbar, A.D. (2015b) Selenium isotopes support free O-2 in the latest Archean. Geology 43, 259-262.
Show in context This suggests that significant free O2 was present in the oceans at that time, which is also supported by several inorganic palaeoredox proxies (Anbar et al., 2007; Reinhard et al., 2009; Stueken et al., 2015b).
View in article
Stueken, E.E., Kipp, M.A., Koehler, M.C., Buick, R. (2016) The evolution of Earth's biogeochemical nitrogen cycle. Earth-Science Reviews 160, 220-239.
Show in context Anammox is not further considered here since its antiquity is still a controversial issue and the resulting δ15NBulk remains unknown (Stueken et al., 2016; see also Supplementary Information).
View in article
Partial NH4+ assimilation can yield ε values down to -27 ‰ when NH4+ concentration is higher than 20 μM, while lower NH4+ concentrations are associated with lower ε values close to -4 ‰ (see Stueken et al., 2016 and references therein).
View in article
Sugitani, K., Grey, K., Allwood, A., Nagaoka, T., Mimura, K., Minami, M., Marshall, C.P., Van Kranendonk, M.J., Walter, M.R. (2007) Diverse microstructures from Archaean chert from the mount goldsworthy-mount grant area, pilbara craton, western australia: Microfossils, dubiofossils, or pseudofossils? Precambrian Research 158, 228-262.
Show in context To investigate this issue, we have characterised the N isotope composition of organic microfossils (noted δ15Nµm) isolated from the mid-Archean Farrel Quartzite formation (3.0 Ga; Sugitani et al., 2007) using nanoscale secondary ion mass spectrometry (NanoSIMS).
View in article
In the acid maceration residue, we identified lenticular, spheroid- and film-like microfossils following the previously reported taxonomy (Sugitani et al., 2007; see also Supplementary Information).
View in article
These microfossils are interpreted as remnants of pelagic microorganisms based on both their morphology (Sugitani et al., 2007; Oehler et al., 2010) and their carbon isotope composition (House et al., 2013).
View in article
Thomazo, C., Ader, M., Philippot, P. (2011) Extreme 15N-enrichments in 2.72-Gyr-old sediments: evidence for a turning point in the nitrogen cycle. Geobiology 9, 107-120.
Show in context The isotopic composition of nitrogen (N) has been investigated in order to tackle questions related to Archean life (Beaumont and Robert, 1999; Pinti et al., 2001; Thomazo et al., 2011; Stueken et al., 2015a).
View in article
Thus, only the partial consumption of NH4+ seems to be an acceptable mechanism to explain the large variation in δ15Nµm values, consistent with an Archean DIN marine reservoir dominated by NH4+ (Thomazo et al., 2011).
View in article
Wang, J.L., Yang, N. (2004) Partial nitrification under limited dissolved oxygen conditions. Process Biochemistry 39, 1223-1229.
Show in context Since aerobic NH4+ oxidation implies the occurrence of free dissolved molecular O2 in the water column (Wang and Yang, 2004), this finding echoes previous geochemical arguments pointing toward mid-Archean oxygen oases and, therefore, biological O2 production by oxygenic photosynthesis in stratified mid-Archean oceans (Crowe et al., 2013; Lyons et al., 2014; Planavsky et al., 2014; Satkoski et al., 2015).
View in article
Westall, F., de Ronde, C.E.J., Southam, G., Grassineau, N., Colas, M., Cockell, C.S., Lammer, H. (2006) Implications of a 3.472-3.333 Gyr-old subaerial microbial mat from the Barberton greenstone belt, South Africa for the UV environmental conditions on the early Earth. Philosophical Transactions of the Royal Society B-Biological Sciences 361, 1857-1875.
Show in context The latter are thus likely remnants of benthic microbial mats, in agreement with their morphology (Westall et al., 2006).
View in article
Supplementary Information
The Supplementary Information includes:
- Studied Samples
- Sample Preparation, Microscopic Observation and Isotopic Analyses
- Biogenicity of the Studied Microfossils
- Reliability of the Morphological Traits and of the Geochemical Composition of Organic-walled Microfossils Isolated by Acid Maceration
- Assessing the Potential Effect of Contamination, Biomass Degradation and Fossilisation on δ15N Values
- Biological Utilisation of NO3- and Anaerobic Ammonia Oxidation (Anammox)
- Tables S-1 and S-2
- Figures S-1 to S-6
- Supplementary Information References
Download the Supplementary Information (PDF).
Figures and Tables
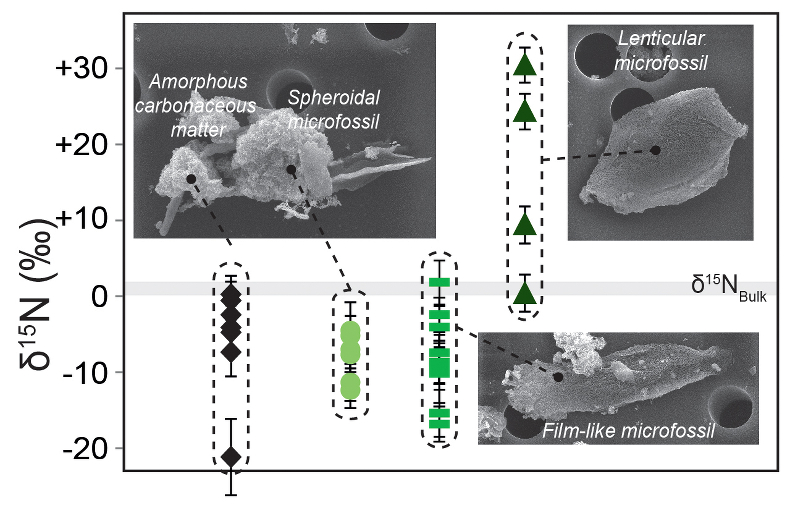
Figure 1 Nitrogen isotopic composition of amorphous carbonaceous matter, and of spheroidal, film-like and lenticular-like microfossils from the 3.0 Ga Farrel Quartzite formation. δ15NBulk values are provided for comparison with δ15Nµm values.
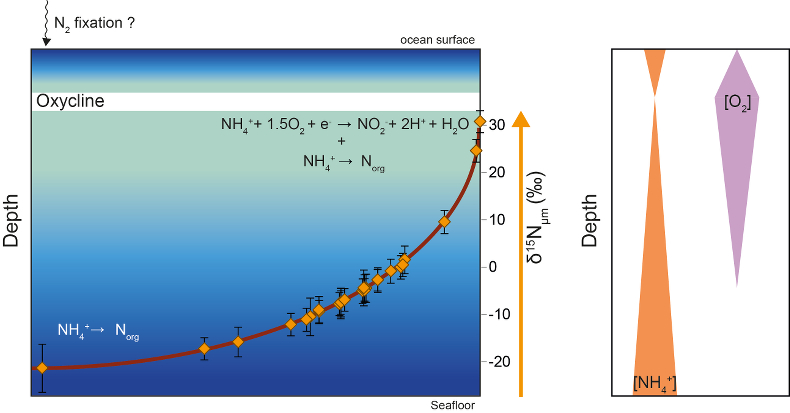
Figure 2 Cartoon illustrating the sedimentary conditions of the Farrel Quartzite formation. The blue gradient in the water column indicates change in NH4+ concentration from high (dark) to low (light) concentrations in anoxic zones. δ15Nµm values determined at the scale of individual microfossil and organic particle are represented by orange diamonds. In the proposed model, negative δ15Nµm values reflect assimilation of NH4+ from benthic efflux whereas positive δ15Nµm values reflect 15N Rayleigh distillation of NH4+ upward through ammonia oxidation when NH4+ concentrations were too low to allow the isotopic fractionation of ammonia assimilation to be expressed (see main text).