



Chemical nature of the 3.4 Ga Strelley Pool microfossils
Affiliations | Corresponding Author | Cite as | Funding informationPublished by the European Association of Geochemistry
under Creative Commons License CC BY 4.0
Keywords: organic microfossils, early life, biosignatures, fossilisation, XANES

- Share this article





Article views:10,611Cumulative count of HTML views and PDF downloads.
- Download Citation
- Rights & Permissions
Associated Press Release

top
Abstract

Figures and Tables
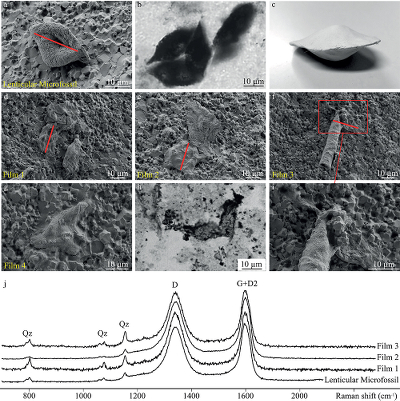 Figure 1 Optical photomicrographs, SEM images and Raman spectra of the 3.4 Ga Strelley Pool organic microfossils observed on a freshly fractured chert fragment. (a) SEM image of a lenticular Strelley Pool microfossil, (b) optical photomicrograph of Strelley Pool lenticular microfossils (Sugitani et al., 2013) and (c) model morphological reconstruction in clay. (d–g) SEM images of Strelley Pool film-like microfossils, (h) optical photomicrograph of a film-like Strelley Pool microfossil (Sugitani et al., 2013) and (i) an enlargement of the film-like microfossil shown in f that looks like a tubular structure. Red lines indicate where the FIB sections were extracted for TEM and STXM analyses (Film 1–4). (j) Raman spectra of the Strelley Pool organic microfossils These spectra are typical of disordered carbon having experienced 300 °C, with the D and composite G + D2 bands displaying similar intensities. Qz corresponds to quartz. | 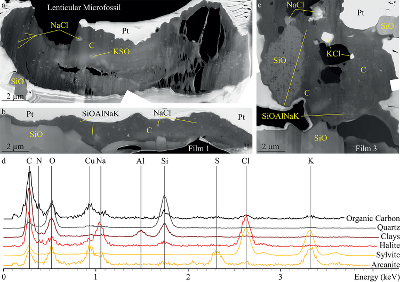 Figure 2 TEM analyses of the Strelley Pool organic microfossils. STEM image of the lenticular microfossil (a) shown in Figure 1a and of the two film-like microfossils (b–c) shown in Figure 1d and 1f, respectively. Organic carbon (which appears darker than mineral phases) is closely associated with micro-quartz, clays and crystals of chlorides and sulphates. The main chemical elements are indicated. (d) TEM-EDX spectra of the organic microfossils and the associated mineral phases. | 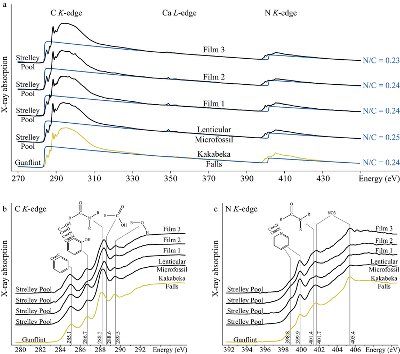 Figure 3 X-ray absorption analyses of the Strelley Pool organic microfossils shown in Figures 1 and 2 and corresponding N/C values. (a) X-ray absorption near edge structure (XANES) spectra at the carbon K-edge and (b) nitrogen K-edge. (c) Diagnostic absorption features include: 285.1 eV for aromatic/olefinic groups, 286.7 eV for imine/nitrile/carbonyl/phenol groups, 288.2 eV for amide groups, 288.6 eV for carboxyl/ester/acetal groups, 289.4 eV for hydroxyl groups, 398.8–399.9 eV - imine/nitrile/aromatic groups, 401.4 eV for amide groups, 401.7 and 405.4 eV for potassium and/or calcium nitrates. A spectrum of a 1.9 Ga Gunflint microfossil (Alleon et al., 2016a) is shown for comparison. |
| Figure 1 | Figure 2 | Figure 3 |
top
Letter
Recent studies have suggested that the biosphere had emerged approximately 4.0 billion years ago (Bell et al., 2015
Bell, E.A., Boehnke, P., Harrison, T.M., Mao, W.L. (2015) Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon. Proceedings of the National Academy of Sciences 112, 14518-14521.
; Dodd et al., 2017Dodd, M.S., Papineau, D., Grenne, T., Slack, J.F., Rittner, M., Pirajno, F., O’Neil, J., Little, C.T. (2017) Evidence for early life in Earth’s oldest hydrothermal vent precipitates. Nature 543, 60-64.
), proliferating and evolving to a ‘sophisticated’ state by 3.7 Ga (Nutman et al., 2016Nutman, A.P., Bennett, V.C., Friend, C.R., Van Kranendonk, M.J., Chivas, A.R. (2016) Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures. Nature 537, 535-538.
; Hassenkam et al., 2017Hassenkam, T., Andersson, M., Dalby, K., Mackenzie, D., Rosing, M. (2017) Elements of Eoarchean life trapped in mineral inclusions. Nature 548, 78-81.
). These conclusions, however, are based on equivocal lines of evidence (e.g., 13C-depleted carbonaceous materials and/or mineral structures) that equally could have been the product of abiotic processes (Grotzinger and Rothman, 1996Grotzinger, J.P., Rothman, D.H. (1996) An abiotic model for stromatolite morphogenesis. Nature 383, 423.
; Garcia-Ruiz et al., 2003Garcia-Ruiz, J., Hyde, S., Carnerup, A., Christy, A., Van Kranendonk, M., Welham, N. (2003) Self-assembled silica-carbonate structures and detection of ancient microfossils. Science 302, 1194-1197.
; McCollom and Seewald, 2006McCollom, T.M., Seewald, J.S. (2006) Carbon isotope composition of organic compounds produced by abiotic synthesis under hydrothermal conditions. Earth and Planetary Science Letters 243, 74-84.
). The absence of indisputable traces of life in Earth’s earliest rocks, specifically the paucity of authentic organic microfossils with biogenically and geologically consistent molecular structures, has prevented the community from reaching a consensus.Unambiguous identification of organic microfossils in the early geological record has always been challenging (Schopf, 1975
Schopf, J.W. (1975) Precambrian paleobiology: problems and perspectives. Annual Review of Earth and Planetary Sciences 3, 213-249.
; Knoll, 2015Knoll, A.H. (2015) Paleobiological perspectives on early microbial evolution. Cold Spring Harbor Perspectives in Biology 7, a018093.
). Quite illustrative is the long-standing debate about the origin of filamentous microstructures from the 3.5 Ga cherts of the Apex Basalt Formation (Pilbara Craton, Australia). There, the disputed microstructures have been interpreted as either remnants of Earth’s oldest cellular life (Schopf and Packer, 1987Schopf, J.W., Packer, B.M. (1987) Early Archean (3.3-billion to 3.5-billion-year-old) microfossils from Warrawoona Group, Australia. Science 237, 70-73.
; Ueno et al., 2001Ueno, Y., Isozaki, Y., Yurimoto, H., Maruyama, S. (2001) Carbon isotopic signatures of individual Archean microfossils (?) from Western Australia. International Geology Review 43, 196-212.
; Schopf et al., 2002Schopf, J.W., Kudryavtsev, A.B., Agresti, D.G., Wdowiak, T.J., Czaja, A.D. (2002) Laser–Raman imagery of Earth's earliest fossils. Nature 416, 73-76.
, 2018Schopf, J.W., Kitajima, K., Spicuzza, M.J., Kudryavtsev, A.B., Valley, J.W. (2018) SIMS analyses of the oldest known assemblage of microfossils document their taxon-correlated carbon isotope compositions. Proceedings of the National Academy of Sciences 115, 53-58.
) or as abiotic organo-mineral structures (Brasier et al., 2002Brasier, M.D., Green, O.R., Jephcoat, A.P., Kleppe, A.K., Van Kranendonk, M.J., Lindsay, J.F., Steele, A., Grassineau, N.V. (2002) Questioning the evidence for Earth's oldest fossils. Nature 416, 76-81.
; Marshall et al., 2011Marshall, C.P., Emry, J.R., Marshall, A.O. (2011) Haematite pseudomicrofossils present in the 3.5-billion-year-old Apex Chert. Nature Geoscience 4, 240.
; Wacey et al., 2016Wacey, D., Saunders, M., Kong, C., Brasier, A., Brasier, M. (2016) 3.46 Ga Apex chert ‘microfossils’ reinterpreted as mineral artefacts produced during phyllosilicate exfoliation. Gondwana Research 36, 296-313.
). Isotopic investigation aside (Ueno et al., 2001Ueno, Y., Isozaki, Y., Yurimoto, H., Maruyama, S. (2001) Carbon isotopic signatures of individual Archean microfossils (?) from Western Australia. International Geology Review 43, 196-212.
; Schopf et al., 2018Schopf, J.W., Kitajima, K., Spicuzza, M.J., Kudryavtsev, A.B., Valley, J.W. (2018) SIMS analyses of the oldest known assemblage of microfossils document their taxon-correlated carbon isotope compositions. Proceedings of the National Academy of Sciences 115, 53-58.
), molecular appraisal of the biogenicity of the Apex microstructures has proven inconclusive, hindered because the analyses were performed on bulk powdered samples rather than targeting the microstructures themselves (De Gregorio et al., 2009De Gregorio, B.T., Sharp, T.G., Flynn, G.J., Wirick, S., Hervig, R.L. (2009) Biogenic origin for Earth's oldest putative microfossils. Geology 37, 631-634.
).To date, the oldest commonly accepted organic microfossils were found within the sedimentary cherts of the 3.4 Ga Strelley Pool Formation from the Panorama greenstone belt (Pilbara Craton, Australia). These cherts are thought to have formed through the crystallisation of colloidal silica that was deposited in a shallow water marine environment, with some hydrothermal inputs (Sugitani et al., 2013
Sugitani, K., Mimura, K., Nagaoka, T., Lepot, K., Takeuchi, M. (2013) Microfossil assemblage from the 3400Ma Strelley Pool Formation in the Pilbara Craton, Western Australia: Results form a new locality. Precambrian Research 226, 59-74.
). Since their formation, these cherts have only undergone low grade metamorphism (prehnite-pumpellyite to lower greenschist facies; Lepot et al., 2013Lepot, K., Williford, K.H., Ushikubo, T., Sugitani, K., Mimura, K., Spicuzza, M.J., Valley, J.W. (2013) Texture-specific isotopic compositions in 3.4 Gyr old organic matter support selective preservation in cell-like structures. Geochimica et Cosmochimica Acta 112, 66-86.
; Sugitani et al., 2013Sugitani, K., Mimura, K., Nagaoka, T., Lepot, K., Takeuchi, M. (2013) Microfossil assemblage from the 3400Ma Strelley Pool Formation in the Pilbara Craton, Western Australia: Results form a new locality. Precambrian Research 226, 59-74.
).The Strelley Pool microfossils display morphologically diverse microstructures (Wacey et al., 2011a
Wacey, D., Kilburn, M.R., Saunders, M., Cliff, J., Brasier, M.D. (2011a) Microfossils of sulphur-metabolizing cells in 3.4-billion-year-old rocks of Western Australia. Nature Geoscience 4, 698-702.
; Sugitani et al., 2013Sugitani, K., Mimura, K., Nagaoka, T., Lepot, K., Takeuchi, M. (2013) Microfossil assemblage from the 3400Ma Strelley Pool Formation in the Pilbara Craton, Western Australia: Results form a new locality. Precambrian Research 226, 59-74.
), featuring 13C-depleted carbon (Lepot et al., 2013Lepot, K., Williford, K.H., Ushikubo, T., Sugitani, K., Mimura, K., Spicuzza, M.J., Valley, J.W. (2013) Texture-specific isotopic compositions in 3.4 Gyr old organic matter support selective preservation in cell-like structures. Geochimica et Cosmochimica Acta 112, 66-86.
), and are sometimes associated with pyrites that exhibit sulphur isotope signatures consistent with a biological origin (Wacey et al., 2011bWacey, D., Saunders, M., Brasier, M.D., Kilburn, M.R. (2011b) Earliest microbially mediated pyrite oxidation in ~3.4 billion-year-old sediments. Earth and Planetary Science Letters 301, 393-402.
). Yet, as for the Apex cherts cited above, the only existing molecular insight into the Strelley Pool organic matter was obtained from powdered material rather than directly from the microfossils (De Gregorio et al., 2011De Gregorio, B.T., Sharp, T.G., Rushdi, A.I., Simoneit, B.R. (2011) Bugs or gunk? Nanoscale methods for assessing the biogenicity of ancient microfossils and organic matter. In: Golding S., Glikson M. (Eds.) Earliest Life on Earth: Habitats, Environments and Methods of Detection. Springer, Dordrecht, 239-289.
).Here, following the methodology adopted for earlier studies (Benzerara et al., 2005
Benzerara, K., Menguy, N., Guyot, F., Vanni, C., Gillet, P. (2005) TEM study of a silicate-carbonate-microbe interface prepared by focused ion beam milling. Geochimica et Cosmochimica Acta 69, 1413-1422.
; Bernard et al., 2007Bernard, S., Benzerara, K., Beyssac, O., Menguy, N., Guyot, F., Brown, G.E., Goffé, B. (2007) Exceptional preservation of fossil plant spores in high-pressure metamorphic rocks. Earth and Planetary Science Letters 262, 257-272.
; Lepot et al., 2008Lepot, K., Benzerara, K., Brown Jr, G.E., Philippot, P. (2008) Microbially influenced formation of 2,724-million-year-old stromatolites. Nature Geoscience 1, 118.
), we exploited advanced microscopy and spectroscopy techniques including Raman microspectroscopy, focused ion beam (FIB), scanning and transmission electron microscopy (SEM and TEM) and scanning transmission X-ray microscopy (STXM) coupled with X-ray absorption near edge structure (XANES) spectroscopy, to conduct spatially‑resolved molecular investigations of the Strelley Pool microfossils at the submicrometre scale. N/C atomic ratio values were estimated following the methodology and calibration outlined in Alleon et al. (2015)Alleon, J., Bernard, S., Remusat, L., Robert, F. (2015) Estimation of nitrogen-to-carbon ratios of organics and carbon materials at the submicrometer scale. Carbon 84, 290-298.
and successfully used in Alleon et al. (2016aAlleon, J., Bernard, S., Le Guillou, C., Marin-Carbonne, J., Pont, S., Beyssac, O., McKeegan, K.D., Robert, F. (2016a) Molecular preservation of 1.88 Ga Gunflint organic microfossils as a function of temperature and mineralogy. Nature Communications 7, 11977.
, 2017Alleon, J., Bernard, S., Le Guillou, C., Daval, D., Skouri-Panet, F., Kuga, M., Robert, F. (2017) Organic molecular heterogeneities can withstand diagenesis. Scientific Reports 7, 1508.
).Consistent with previous observations of thin sections (Lepot et al., 2013
Lepot, K., Williford, K.H., Ushikubo, T., Sugitani, K., Mimura, K., Spicuzza, M.J., Valley, J.W. (2013) Texture-specific isotopic compositions in 3.4 Gyr old organic matter support selective preservation in cell-like structures. Geochimica et Cosmochimica Acta 112, 66-86.
; Sugitani et al., 2013Sugitani, K., Mimura, K., Nagaoka, T., Lepot, K., Takeuchi, M. (2013) Microfossil assemblage from the 3400Ma Strelley Pool Formation in the Pilbara Craton, Western Australia: Results form a new locality. Precambrian Research 226, 59-74.
; Fig. 1b, h), SEM observations reveal the presence of lenticular and film-like organic microfossils present on freshly fractured faces of the PAN1-1A chert sample (Fig. 1d–g, i). Raman data collected from each of the individual microfossils demonstrate the poorly ordered nature of the organic material that typifies these microfossils (Fig. 1j). Application of the RSCM (Raman Spectroscopy of Carbonaceous Material) geothermometer (Beyssac et al., 2002Beyssac, O., Goffé, B., Chopin, C., Rouzaud, J. (2002) Raman spectra of carbonaceous material in metasediments: a new geothermometer. Journal of Metamorphic Geology 20, 859-871.
; Lahfid et al., 2010Lahfid, A., Beyssac, O., Deville, E., Negro, F., Chopin, C., Goffé, B. (2010) Evolution of the Raman spectrum of carbonaceous material in low‐grade metasediments of the Glarus Alps (Switzerland). Terra Nova 22, 354-360.
) indicates that the Strelley Pool microfossils experienced peak temperatures of approximately 300 °C, which is in good agreement with regional metamorphic histories (Wacey et al., 2011aWacey, D., Kilburn, M.R., Saunders, M., Cliff, J., Brasier, M.D. (2011a) Microfossils of sulphur-metabolizing cells in 3.4-billion-year-old rocks of Western Australia. Nature Geoscience 4, 698-702.
; Lepot et al., 2013Lepot, K., Williford, K.H., Ushikubo, T., Sugitani, K., Mimura, K., Spicuzza, M.J., Valley, J.W. (2013) Texture-specific isotopic compositions in 3.4 Gyr old organic matter support selective preservation in cell-like structures. Geochimica et Cosmochimica Acta 112, 66-86.
; Sugitani et al., 2013Sugitani, K., Mimura, K., Nagaoka, T., Lepot, K., Takeuchi, M. (2013) Microfossil assemblage from the 3400Ma Strelley Pool Formation in the Pilbara Craton, Western Australia: Results form a new locality. Precambrian Research 226, 59-74.
). Transmission electron microscopy conducted on FIB foils demonstrates that, even at the nanometre scale, the organic matter is intimately associated with quartz and clays, as well as crystalline chlorides or sulphate minerals (Fig. 2). Assimilated, these observations confirm the syngenicity of the Strelley Pool microfossils, testifying that they were entombed within the chert at the time of its formation.Figure 1 Optical photomicrographs, SEM images and Raman spectra of the 3.4 Ga Strelley Pool organic microfossils observed on a freshly fractured chert fragment. (a) SEM image of a lenticular Strelley Pool microfossil, (b) optical photomicrograph of Strelley Pool lenticular microfossils (Sugitani et al., 2013
Sugitani, K., Mimura, K., Nagaoka, T., Lepot, K., Takeuchi, M. (2013) Microfossil assemblage from the 3400Ma Strelley Pool Formation in the Pilbara Craton, Western Australia: Results form a new locality. Precambrian Research 226, 59-74.
) and (c) model morphological reconstruction in clay. (d–g) SEM images of Strelley Pool film-like microfossils, (h) optical photomicrograph of a film-like Strelley Pool microfossil (Sugitani et al., 2013Sugitani, K., Mimura, K., Nagaoka, T., Lepot, K., Takeuchi, M. (2013) Microfossil assemblage from the 3400Ma Strelley Pool Formation in the Pilbara Craton, Western Australia: Results form a new locality. Precambrian Research 226, 59-74.
) and (i) an enlargement of the film-like microfossil shown in f that looks like a tubular structure. Red lines indicate where the FIB sections were extracted for TEM and STXM analyses (Film 1–4). (j) Raman spectra of the Strelley Pool organic microfossils These spectra are typical of disordered carbon having experienced 300 °C, with the D and composite G + D2 bands displaying similar intensities. Qz corresponds to quartz.Figure 2 TEM analyses of the Strelley Pool organic microfossils. STEM image of the lenticular microfossil (a) shown in Figure 1a and of the two film-like microfossils (b–c) shown in Figure 1d and 1f, respectively. Organic carbon (which appears darker than mineral phases) is closely associated with micro-quartz, clays and crystals of chlorides and sulphates. The main chemical elements are indicated. (d) TEM-EDX spectra of the organic microfossils and the associated mineral phases.
Spatially-resolved chemical investigation exploiting STXM reveal that the Strelley Pool microfossils possess very high N/C values (0.24 ± 0.02; Fig. 3a). Although the microfossils are associated with potassium and calcium nitrates, these phases are only present in trace amounts and do not significantly contribute to the estimated N/C values (cf. Supplementary Information). The XANES spectra of the Strelley Pool microfossils resemble those reported from the best preserved microfossils found in the 1.9 Ga Gunflint cherts (Fig. 3b–c; Alleon et al., 2016a
Alleon, J., Bernard, S., Le Guillou, C., Marin-Carbonne, J., Pont, S., Beyssac, O., McKeegan, K.D., Robert, F. (2016a) Molecular preservation of 1.88 Ga Gunflint organic microfossils as a function of temperature and mineralogy. Nature Communications 7, 11977.
). Here, the C-XANES spectra display multiple peaks: the intense peak at energies ranging between 288.2 and 288.6 eV, likely reflects a mixed contribution from amide ((R1, R2)N–C=O), carboxylic (COOH), ester (COOR) and/or acetal (C(OR)2) groups. The peak at 289.5 eV signals the presence of hydroxyl (OH) groups. The peak at 286.7 eV reveals the presence of imine (C=N), nitrile (C≡N), carbonyl (C=O) and/or phenolic (Ar–OH) groups. Whereas, the much smaller peak centred at 285.1 eV signals the presence of aromatic and/or olefinic groups (De Gregorio et al., 2011De Gregorio, B.T., Sharp, T.G., Rushdi, A.I., Simoneit, B.R. (2011) Bugs or gunk? Nanoscale methods for assessing the biogenicity of ancient microfossils and organic matter. In: Golding S., Glikson M. (Eds.) Earliest Life on Earth: Habitats, Environments and Methods of Detection. Springer, Dordrecht, 239-289.
; Alleon et al., 2017Alleon, J., Bernard, S., Le Guillou, C., Daval, D., Skouri-Panet, F., Kuga, M., Robert, F. (2017) Organic molecular heterogeneities can withstand diagenesis. Scientific Reports 7, 1508.
). Similarly, the N-XANES spectra display a broad absorption feature at 401.4 eV, which is related to the presence of amide, imine and/or nitrile groups. The peak at 399.9 eV and shoulder around 398.8 eV indicate the presence of nitrogen within aromatic moieties (Leinweber et al., 2007Leinweber, P., Kruse, J., Walley, F.L., Gillespie, A., Eckhardt, K.-U., Blyth, R.I., Regier, T. (2007) Nitrogen K-edge XANES–an overview of reference compounds used to identify unknown organic nitrogen in environmental samples. Journal of Synchrotron Radiation 14, 500-511.
; De Gregorio et al., 2011De Gregorio, B.T., Sharp, T.G., Rushdi, A.I., Simoneit, B.R. (2011) Bugs or gunk? Nanoscale methods for assessing the biogenicity of ancient microfossils and organic matter. In: Golding S., Glikson M. (Eds.) Earliest Life on Earth: Habitats, Environments and Methods of Detection. Springer, Dordrecht, 239-289.
; Alleon et al., 2017Alleon, J., Bernard, S., Le Guillou, C., Daval, D., Skouri-Panet, F., Kuga, M., Robert, F. (2017) Organic molecular heterogeneities can withstand diagenesis. Scientific Reports 7, 1508.
). The two absorption features at 401.7 and 405.4 eV can be attributed to the presence of trace quantities of potassium and/or calcium nitrates (Leinweber et al., 2007Leinweber, P., Kruse, J., Walley, F.L., Gillespie, A., Eckhardt, K.-U., Blyth, R.I., Regier, T. (2007) Nitrogen K-edge XANES–an overview of reference compounds used to identify unknown organic nitrogen in environmental samples. Journal of Synchrotron Radiation 14, 500-511.
; Alleon et al., 2016aAlleon, J., Bernard, S., Le Guillou, C., Marin-Carbonne, J., Pont, S., Beyssac, O., McKeegan, K.D., Robert, F. (2016a) Molecular preservation of 1.88 Ga Gunflint organic microfossils as a function of temperature and mineralogy. Nature Communications 7, 11977.
). Altogether, consistent with previous claims (Lepot et al., 2013Lepot, K., Williford, K.H., Ushikubo, T., Sugitani, K., Mimura, K., Spicuzza, M.J., Valley, J.W. (2013) Texture-specific isotopic compositions in 3.4 Gyr old organic matter support selective preservation in cell-like structures. Geochimica et Cosmochimica Acta 112, 66-86.
; Sugitani et al., 2013Sugitani, K., Mimura, K., Nagaoka, T., Lepot, K., Takeuchi, M. (2013) Microfossil assemblage from the 3400Ma Strelley Pool Formation in the Pilbara Craton, Western Australia: Results form a new locality. Precambrian Research 226, 59-74.
), the spatially resolved chemical information reported here supports the biogenicity of the Strelley Pool microfossils, advocating them as the oldest currently known authentic microfossils.Figure 3 X-ray absorption analyses of the Strelley Pool organic microfossils shown in Figures 1 and 2 and corresponding N/C values. (a) X-ray absorption near edge structure (XANES) spectra at the carbon K-edge and (b) nitrogen K-edge. (c) Diagnostic absorption features include: 285.1 eV for aromatic/olefinic groups, 286.7 eV for imine/nitrile/carbonyl/phenol groups, 288.2 eV for amide groups, 288.6 eV for carboxyl/ester/acetal groups, 289.4 eV for hydroxyl groups, 398.8–399.9 eV - imine/nitrile/aromatic groups, 401.4 eV for amide groups, 401.7 and 405.4 eV for potassium and/or calcium nitrates. A spectrum of a 1.9 Ga Gunflint microfossil (Alleon et al., 2016a
Alleon, J., Bernard, S., Le Guillou, C., Marin-Carbonne, J., Pont, S., Beyssac, O., McKeegan, K.D., Robert, F. (2016a) Molecular preservation of 1.88 Ga Gunflint organic microfossils as a function of temperature and mineralogy. Nature Communications 7, 11977.
) is shown for comparison.The general palaeobiological perception is that organic diagenesis results in a convergence in the composition of organic materials from different sources, reducing the utility of chemical composition for discriminating between taxa (Briggs and McMahon, 2016
Briggs, D.E., McMahon, S. (2016) The role of experiments in investigating the taphonomy of exceptional preservation. Palaeontology 59, 1-11.
). Yet, the lenticular and film-like Strelley Pool microfossils do not produce identical XANES spectra. The film-like microfossils contain slightly more abundant carboxylic, ester and/or acetal groups and fewer amide groups than their lenticular counterpart (Fig. 3). Recent laboratory experiments have demonstrated that thermal degradation of chemically distinct organic precursors can lead to chemically distinct residues (Alleon et al., 2017Alleon, J., Bernard, S., Le Guillou, C., Daval, D., Skouri-Panet, F., Kuga, M., Robert, F. (2017) Organic molecular heterogeneities can withstand diagenesis. Scientific Reports 7, 1508.
). The spectral differences between the lenticular and film-like microfossils may thus signal differences in the initial molecular composition of organic material now composing the Strelley Pool microfossils. The data reported here therefore suggest inheritance from slight, yet morphotype-specific, differences in the initial proportions of proteins, sugars and lipids of the Strelley Pool microfossils.The Strelley Pool microfossils investigated here exhibit high N/C values and contain amide functional groups despite a 3.4 Gyr geological history and heating episodes up to 300 °C, i.e. conditions that normally lead to intense chemical degradation (Bernard and Horsfield, 2014
Bernard, S., Horsfield, B. (2014) Thermal maturation of gas shale systems. Annual Review of Earth and Planetary Sciences 42, 635-651.
; Briggs and Summons, 2014Briggs, D.E., Summons, R.E. (2014) Ancient biomolecules: their origins, fossilization, and role in revealing the history of life. BioEssays 36, 482-490.
). Such a high level of molecular preservation was achieved likely because of their early silicification. A similar mechanism was proposed for the chemically well preserved 1.9 Ga Gunflint microfossils that underwent temperatures up to 150–170 °C (Alleon et al., 2016aAlleon, J., Bernard, S., Le Guillou, C., Marin-Carbonne, J., Pont, S., Beyssac, O., McKeegan, K.D., Robert, F. (2016a) Molecular preservation of 1.88 Ga Gunflint organic microfossils as a function of temperature and mineralogy. Nature Communications 7, 11977.
). Consistently, recent laboratory experiments demonstrated that early entombment in colloidal silica significantly reduces thermally induced molecular degradation of microorganisms, even if the silica ends up being completely converted into thermally stable quartz (Alleon et al., 2016bAlleon, J., Bernard, S., Le Guillou, C., Daval, D., Skouri-Panet, F., Pont, S., Delbes, L., Robert, F. (2016b) Early entombment within silica minimizes the molecular degradation of microorganisms during advanced diagenesis. Chemical Geology 437, 98-108.
).Surprisingly, the 3.4 Ga Strelley Pool microfossils are better preserved than the 1.9 Ga Gunflint microfossils having experienced temperatures of about 200-220 °C (Alleon et al., 2016a
Alleon, J., Bernard, S., Le Guillou, C., Marin-Carbonne, J., Pont, S., Beyssac, O., McKeegan, K.D., Robert, F. (2016a) Molecular preservation of 1.88 Ga Gunflint organic microfossils as a function of temperature and mineralogy. Nature Communications 7, 11977.
). These chemically more degraded Gunflint microfossils have much lower N/C values and do not contain any detectable amide groups (Alleon et al., 2016aAlleon, J., Bernard, S., Le Guillou, C., Marin-Carbonne, J., Pont, S., Beyssac, O., McKeegan, K.D., Robert, F. (2016a) Molecular preservation of 1.88 Ga Gunflint organic microfossils as a function of temperature and mineralogy. Nature Communications 7, 11977.
). This difference may be related to differences in porosity of the chert matrices. Cherts have essentially no measurable permeability (Knauth, 2005Knauth, L.P. (2005) Temperature and salinity history of the Precambrian ocean: implications for the course of microbial evolution. Palaeogeography, Palaeoclimatology, Palaeoecology 219, 53-69.
), which generally excludes the circulation of oxygenated diagenetic fluids which would impair molecular information. This was the case for the Strelley Pool chert, as supported by the absence of secondary minerals, but not for the most hydrothermally altered Gunflint cherts, as evidenced by the presence of iron oxides that likely formed during diagenesis (Shapiro and Konhauser, 2015Shapiro, R., Konhauser, K. (2015) Hematite‐coated microfossils: primary ecological fingerprint or taphonomic oddity of the Paleoproterozoic? Geobiology 13, 209-224.
; Alleon et al., 2016aAlleon, J., Bernard, S., Le Guillou, C., Marin-Carbonne, J., Pont, S., Beyssac, O., McKeegan, K.D., Robert, F. (2016a) Molecular preservation of 1.88 Ga Gunflint organic microfossils as a function of temperature and mineralogy. Nature Communications 7, 11977.
). Assimilating these observations, we speculate that timing and severity of permeability loss may be more critical than temperature for the molecular preservation of organic microfossils.The present study demonstrates that ancient organic microfossils may exhibit a high level of chemical preservation in appropriate settings independent of a long and complex geological history. These findings highlight the potential of the oldest geological record to offer a unique window into the Earth’s juvenile biosphere and, eventually, to place constraints on the timing, tempo and trajectory of its evolution.
top
Acknowledgements
We gratefully acknowledge financial support from the ERC (project PaleoNanoLife—PI: FR). We thank Mélinée Deretz for administrative support, Imène Esteve (IMPMC) for support with the SEM, David Troadec (IEMN) for the preparation of FIB sections, Jian Wang and Yingshen Lu for support with the STXM at the Canadian Light Source (CLS) and to Stefan Stanescu and Sufal Swaraj for support with the STXM at SOLEIL. Kevin Lepot (LOG) is acknowledged for formative discussions. JA acknowledges the Simons Foundation under the auspices of the Simons Collaboration on the Origins of Life award #290361 for support during the preparation of this manuscript. We thank the 4 anonymous reviewers for their constructive and helpful comments. Gareth Izon (MIT) read the manuscript and clarified some of the text. The SEM facility of the IMPMC is supported by Region Ile de France grant SESAME Number I-07-593/R, INSU-CNRS, INP-CNRS and UPMC-Paris 6, and by the Agence Nationale de la Recherche (ANR) grant number ANR-07-BLAN-0124-01. The TEM facility at the University of Lille is supported by the Chevreul Institute, the European FEDER and Région Nord-Pas-de-Calais. STXM-based X-ray absorption spectroscopy data were acquired at the 10ID-1 beamline at the CLS, which is supported by the NSERC, the CIHR, the NRC and the University of Saskatchewan and at the HERMES beamline at SOLEIL, which is supported by the CNRS, the CEA, the Région Ile de France, the Departmental Council of Essonne and the Région Centre.
Editor: Liane G. Benning
top
Author Contributions
JA, SB and FR conceived and designed the research. JA and SB performed the SEM experiments. JA, SB and OB performed the Raman spectroscopy experiments. CLG. performed the TEM experiments. JA, SB and CLG performed the STXM-based XANES experiments, interpreted the data and wrote the manuscript with contributions from OB, KS and FR.
top
References
Alleon, J., Bernard, S., Remusat, L., Robert, F. (2015) Estimation of nitrogen-to-carbon ratios of organics and carbon materials at the submicrometer scale. Carbon 84, 290-298.
 Show in context
Show in context N/C atomic ratio values were estimated following the methodology and calibration outlined in Alleon et al. (2015) and successfully used in Alleon et al. (2016a, 2017).
View in article
Alleon, J., Bernard, S., Le Guillou, C., Marin-Carbonne, J., Pont, S., Beyssac, O., McKeegan, K.D., Robert, F. (2016a) Molecular preservation of 1.88 Ga Gunflint organic microfossils as a function of temperature and mineralogy. Nature Communications 7, 11977.
Show in context N/C atomic ratio values were estimated following the methodology and calibration outlined in Alleon et al. (2015) and successfully used in Alleon et al. (2016a, 2017).
View in article
The XANES spectra of the Strelley Pool microfossils resemble those reported from the best preserved microfossils found in the 1.9 Ga Gunflint cherts (Fig. 3b–c; Alleon et al., 2016a).
View in article
The two absorption features at 401.7 and 405.4 eV can be attributed to the presence of trace quantities of potassium and/or calcium nitrates (Leinweber et al., 2007; Alleon et al., 2016a).
View in article
Figure 3 [...] A spectrum of a 1.9 Ga Gunflint microfossil (Alleon et al., 2016a) is shown for comparison.
View in article
A similar mechanism was proposed for the chemically well preserved 1.9 Ga Gunflint microfossils that underwent temperatures up to 150–170 °C (Alleon et al., 2016a).
View in article
Surprisingly, the 3.4 Ga Strelley Pool microfossils are better preserved than the 1.9 Ga Gunflint microfossils having experienced temperatures of about 200-220 °C (Alleon et al., 2016a).
View in article
These chemically more degraded Gunflint microfossils have much lower N/C values and do not contain any detectable amide groups (Alleon et al., 2016a).
View in article
This was the case for the Strelley Pool chert, as supported by the absence of secondary minerals, but not for the most hydrothermally altered Gunflint cherts, as evidenced by the presence of iron oxides that likely formed during diagenesis (Shapiro and Konhauser, 2015; Alleon et al., 2016a).
View in article
Alleon, J., Bernard, S., Le Guillou, C., Daval, D., Skouri-Panet, F., Pont, S., Delbes, L., Robert, F. (2016b) Early entombment within silica minimizes the molecular degradation of microorganisms during advanced diagenesis. Chemical Geology 437, 98-108.
Show in context Consistently, recent laboratory experiments demonstrated that early entombment in colloidal silica significantly reduces thermally induced molecular degradation of microorganisms, even if the silica ends up being completely converted into thermally stable quartz (Alleon et al., 2016b).
View in article
Alleon, J., Bernard, S., Le Guillou, C., Daval, D., Skouri-Panet, F., Kuga, M., Robert, F. (2017) Organic molecular heterogeneities can withstand diagenesis. Scientific Reports 7, 1508.
Show in context N/C atomic ratio values were estimated following the methodology and calibration outlined in Alleon et al. (2015) and successfully used in Alleon et al. (2016a, 2017).
View in article
Whereas, the much smaller peak centred at 285.1 eV signals the presence of aromatic and/or olefinic groups (De Gregorio et al., 2011; Alleon et al., 2017).
View in article
The peak at 399.9 eV and shoulder around 398.8 eV indicate the presence of nitrogen within aromatic moieties (Leinweber et al., 2007; De Gregorio et al., 2011; Alleon et al., 2017).
View in article
Recent laboratory experiments have demonstrated that thermal degradation of chemically distinct organic precursors can lead to chemically distinct residues (Alleon et al., 2017).
View in article
Bell, E.A., Boehnke, P., Harrison, T.M., Mao, W.L. (2015) Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon. Proceedings of the National Academy of Sciences 112, 14518-14521.
Show in context Recent studies have suggested that the biosphere had emerged approximately 4.0 billion years ago (Bell et al., 2015; Dodd et al., 2017), proliferating and evolving to a ‘sophisticated’ state by 3.7 Ga (Nutman et al., 2016; Hassenkam et al., 2017).
View in article
Benzerara, K., Menguy, N., Guyot, F., Vanni, C., Gillet, P. (2005) TEM study of a silicate-carbonate-microbe interface prepared by focused ion beam milling. Geochimica et Cosmochimica Acta 69, 1413-1422.
Show in context Here, following the methodology adopted for earlier studies (Benzerara et al., 2005; Bernard et al., 2007; Lepot et al., 2008), we exploited advanced microscopy and spectroscopy techniques including Raman microspectroscopy, focused ion beam (FIB), scanning and transmission electron microscopy (SEM and TEM) and scanning transmission X-ray microscopy (STXM) coupled with X-ray absorption near edge structure (XANES) spectroscopy, to conduct spatially‑resolved molecular investigations of the Strelley Pool microfossils at the submicrometre scale.
View in article
Bernard, S., Benzerara, K., Beyssac, O., Menguy, N., Guyot, F., Brown, G.E., Goffé, B. (2007) Exceptional preservation of fossil plant spores in high-pressure metamorphic rocks. Earth and Planetary Science Letters 262, 257-272.
Show in context Here, following the methodology adopted for earlier studies (Benzerara et al., 2005; Bernard et al., 2007; Lepot et al., 2008), we exploited advanced microscopy and spectroscopy techniques including Raman microspectroscopy, focused ion beam (FIB), scanning and transmission electron microscopy (SEM and TEM) and scanning transmission X-ray microscopy (STXM) coupled with X-ray absorption near edge structure (XANES) spectroscopy, to conduct spatially‑resolved molecular investigations of the Strelley Pool microfossils at the submicrometre scale.
View in article
Bernard, S., Horsfield, B. (2014) Thermal maturation of gas shale systems. Annual Review of Earth and Planetary Sciences 42, 635-651.
Show in context The Strelley Pool microfossils investigated here exhibit high N/C values and contain amide functional groups despite a 3.4 Gyr geological history and heating episodes up to 300 °C, i.e. conditions that normally lead to intense chemical degradation (Bernard and Horsfield, 2014; Briggs and Summons, 2014).
View in article
Beyssac, O., Goffé, B., Chopin, C., Rouzaud, J. (2002) Raman spectra of carbonaceous material in metasediments: a new geothermometer. Journal of Metamorphic Geology 20, 859-871.
Show in context Application of the RSCM (Raman Spectroscopy of Carbonaceous Material) geothermometer (Beyssac et al., 2002; Lahfid et al., 2010) indicates that the Strelley Pool microfossils experienced peak temperatures of approximately 300 °C, which is in good agreement with regional metamorphic histories (Wacey et al., 2011a; Lepot et al., 2013; Sugitani et al., 2013).
View in article
Brasier, M.D., Green, O.R., Jephcoat, A.P., Kleppe, A.K., Van Kranendonk, M.J., Lindsay, J.F., Steele, A., Grassineau, N.V. (2002) Questioning the evidence for Earth's oldest fossils. Nature 416, 76-81.
Show in context There, the disputed microstructures have been interpreted as either remnants of Earth’s oldest cellular life (Schopf and Packer, 1987; Ueno et al., 2001; Schopf et al., 2002, 2018) or as abiotic organo-mineral structures (Brasier et al., 2002; Marshall et al., 2011; Wacey et al., 2016).
View in article
Briggs, D.E., Summons, R.E. (2014) Ancient biomolecules: their origins, fossilization, and role in revealing the history of life. BioEssays 36, 482-490.
Show in context The Strelley Pool microfossils investigated here exhibit high N/C values and contain amide functional groups despite a 3.4 Gyr geological history and heating episodes up to 300 °C, i.e. conditions that normally lead to intense chemical degradation (Bernard and Horsfield, 2014; Briggs and Summons, 2014).
View in article
Briggs, D.E., McMahon, S. (2016) The role of experiments in investigating the taphonomy of exceptional preservation. Palaeontology 59, 1-11.
Show in context The general palaeobiological perception is that organic diagenesis results in a convergence in the composition of organic materials from different sources, reducing the utility of chemical composition for discriminating between taxa (Briggs and McMahon, 2016).
View in article
De Gregorio, B.T., Sharp, T.G., Flynn, G.J., Wirick, S., Hervig, R.L. (2009) Biogenic origin for Earth's oldest putative microfossils. Geology 37, 631-634.
Show in context Isotopic investigation aside (Ueno et al., 2001; Schopf et al., 2018), molecular appraisal of the biogenicity of the Apex microstructures has proven inconclusive, hindered because the analyses were performed on bulk powdered samples rather than targeting the microstructures themselves (De Gregorio et al., 2009).
View in article
De Gregorio, B.T., Sharp, T.G., Rushdi, A.I., Simoneit, B.R. (2011) Bugs or gunk? Nanoscale methods for assessing the biogenicity of ancient microfossils and organic matter. In: Golding S., Glikson M. (Eds.) Earliest Life on Earth: Habitats, Environments and Methods of Detection. Springer, Dordrecht, 239-289.
Show in context Yet, as for the Apex cherts cited above, the only existing molecular insight into the Strelley Pool organic matter was obtained from powdered material rather than directly from the microfossils (De Gregorio et al., 2011).
View in article
Whereas, the much smaller peak centred at 285.1 eV signals the presence of aromatic and/or olefinic groups (De Gregorio et al., 2011; Alleon et al., 2017).
View in article
The peak at 399.9 eV and shoulder around 398.8 eV indicate the presence of nitrogen within aromatic moieties (Leinweber et al., 2007; De Gregorio et al., 2011; Alleon et al., 2017).
View in article
Dodd, M.S., Papineau, D., Grenne, T., Slack, J.F., Rittner, M., Pirajno, F., O’Neil, J., Little, C.T. (2017) Evidence for early life in Earth’s oldest hydrothermal vent precipitates. Nature 543, 60-64.
Show in context Recent studies have suggested that the biosphere had emerged approximately 4.0 billion years ago (Bell et al., 2015; Dodd et al., 2017), proliferating and evolving to a ‘sophisticated’ state by 3.7 Ga (Nutman et al., 2016; Hassenkam et al., 2017).
View in article
Garcia-Ruiz, J., Hyde, S., Carnerup, A., Christy, A., Van Kranendonk, M., Welham, N. (2003) Self-assembled silica-carbonate structures and detection of ancient microfossils. Science 302, 1194-1197.
Show in context These conclusions, however, are based on equivocal lines of evidence (e.g., 13C-depleted carbonaceous materials and/or mineral structures) that equally could have been the product of abiotic processes (Grotzinger and Rothman, 1996; Garcia-Ruiz et al., 2003; McCollom and Seewald, 2006).
View in article
Grotzinger, J.P., Rothman, D.H. (1996) An abiotic model for stromatolite morphogenesis. Nature 383, 423.
Show in context These conclusions, however, are based on equivocal lines of evidence (e.g., 13C-depleted carbonaceous materials and/or mineral structures) that equally could have been the product of abiotic processes (Grotzinger and Rothman, 1996; Garcia-Ruiz et al., 2003; McCollom and Seewald, 2006).
View in article
Hassenkam, T., Andersson, M., Dalby, K., Mackenzie, D., Rosing, M. (2017) Elements of Eoarchean life trapped in mineral inclusions. Nature 548, 78-81.
Show in context Recent studies have suggested that the biosphere had emerged approximately 4.0 billion years ago (Bell et al., 2015; Dodd et al., 2017), proliferating and evolving to a ‘sophisticated’ state by 3.7 Ga (Nutman et al., 2016; Hassenkam et al., 2017).
View in article
Knauth, L.P. (2005) Temperature and salinity history of the Precambrian ocean: implications for the course of microbial evolution. Palaeogeography, Palaeoclimatology, Palaeoecology 219, 53-69.
Show in context Cherts have essentially no measurable permeability (Knauth, 2005), which generally excludes the circulation of oxygenated diagenetic fluids which would impair molecular information.
View in article
Knoll, A.H. (2015) Paleobiological perspectives on early microbial evolution. Cold Spring Harbor Perspectives in Biology 7, a018093.
Show in context Unambiguous identification of organic microfossils in the early geological record has always been challenging (Schopf, 1975; Knoll, 2015).
View in article
Lahfid, A., Beyssac, O., Deville, E., Negro, F., Chopin, C., Goffé, B. (2010) Evolution of the Raman spectrum of carbonaceous material in low‐grade metasediments of the Glarus Alps (Switzerland). Terra Nova 22, 354-360.
Show in context Application of the RSCM (Raman Spectroscopy of Carbonaceous Material) geothermometer (Beyssac et al., 2002; Lahfid et al., 2010) indicates that the Strelley Pool microfossils experienced peak temperatures of approximately 300 °C, which is in good agreement with regional metamorphic histories (Wacey et al., 2011a; Lepot et al., 2013; Sugitani et al., 2013).
View in article
Leinweber, P., Kruse, J., Walley, F.L., Gillespie, A., Eckhardt, K.-U., Blyth, R.I., Regier, T. (2007) Nitrogen K-edge XANES–an overview of reference compounds used to identify unknown organic nitrogen in environmental samples. Journal of Synchrotron Radiation 14, 500-511.
Show in context The peak at 399.9 eV and shoulder around 398.8 eV indicate the presence of nitrogen within aromatic moieties (Leinweber et al., 2007; De Gregorio et al., 2011; Alleon et al., 2017).
View in article
The two absorption features at 401.7 and 405.4 eV can be attributed to the presence of trace quantities of potassium and/or calcium nitrates (Leinweber et al., 2007; Alleon et al., 2016a).
View in article
Lepot, K., Benzerara, K., Brown Jr, G.E., Philippot, P. (2008) Microbially influenced formation of 2,724-million-year-old stromatolites. Nature Geoscience 1, 118.
Show in context Here, following the methodology adopted for earlier studies (Benzerara et al., 2005; Bernard et al., 2007; Lepot et al., 2008), we exploited advanced microscopy and spectroscopy techniques including Raman microspectroscopy, focused ion beam (FIB), scanning and transmission electron microscopy (SEM and TEM) and scanning transmission X-ray microscopy (STXM) coupled with X-ray absorption near edge structure (XANES) spectroscopy, to conduct spatially‑resolved molecular investigations of the Strelley Pool microfossils at the submicrometre scale.
View in article
Lepot, K., Williford, K.H., Ushikubo, T., Sugitani, K., Mimura, K., Spicuzza, M.J., Valley, J.W. (2013) Texture-specific isotopic compositions in 3.4 Gyr old organic matter support selective preservation in cell-like structures. Geochimica et Cosmochimica Acta 112, 66-86.
Show in context Since their formation, these cherts have only undergone low grade metamorphism (prehnite-pumpellyite to lower greenschist facies; Lepot et al., 2013; Sugitani et al., 2013).
View in article
The Strelley Pool microfossils display morphologically diverse microstructures (Wacey et al., 2011a; Sugitani et al., 2013), featuring 13C-depleted carbon (Lepot et al., 2013), and are sometimes associated with pyrites that exhibit sulphur isotope signatures consistent with a biological origin (Wacey et al., 2011b).
View in article
Consistent with previous observations of thin sections (Lepot et al., 2013; Sugitani et al., 2013; Fig. 1b, h), SEM observations reveal the presence of lenticular and film-like organic microfossils present on freshly fractured faces of the PAN1-1A chert sample (Fig. 1d–g, i).
View in article
Application of the RSCM (Raman Spectroscopy of Carbonaceous Material) geothermometer (Beyssac et al., 2002; Lahfid et al., 2010) indicates that the Strelley Pool microfossils experienced peak temperatures of approximately 300 °C, which is in good agreement with regional metamorphic histories (Wacey et al., 2011a; Lepot et al., 2013; Sugitani et al., 2013).
View in article
Altogether, consistent with previous claims (Lepot et al., 2013; Sugitani et al., 2013), the spatially resolved chemical information reported here supports the biogenicity of the Strelley Pool microfossils, advocating them as the oldest currently known authentic microfossils.
View in article
Marshall, C.P., Emry, J.R., Marshall, A.O. (2011) Haematite pseudomicrofossils present in the 3.5-billion-year-old Apex Chert. Nature Geoscience 4, 240.
Show in context There, the disputed microstructures have been interpreted as either remnants of Earth’s oldest cellular life (Schopf and Packer, 1987; Ueno et al., 2001; Schopf et al., 2002, 2018) or as abiotic organo-mineral structures (Brasier et al., 2002; Marshall et al., 2011; Wacey et al., 2016).
View in article
McCollom, T.M., Seewald, J.S. (2006) Carbon isotope composition of organic compounds produced by abiotic synthesis under hydrothermal conditions. Earth and Planetary Science Letters 243, 74-84.
Show in context These conclusions, however, are based on equivocal lines of evidence (e.g., 13C-depleted carbonaceous materials and/or mineral structures) that equally could have been the product of abiotic processes (Grotzinger and Rothman, 1996; Garcia-Ruiz et al., 2003; McCollom and Seewald, 2006).
View in article
Nutman, A.P., Bennett, V.C., Friend, C.R., Van Kranendonk, M.J., Chivas, A.R. (2016) Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures. Nature 537, 535-538.
Show in context Recent studies have suggested that the biosphere had emerged approximately 4.0 billion years ago (Bell et al., 2015; Dodd et al., 2017), proliferating and evolving to a ‘sophisticated’ state by 3.7 Ga (Nutman et al., 2016; Hassenkam et al., 2017).
View in article
Schopf, J.W. (1975) Precambrian paleobiology: problems and perspectives. Annual Review of Earth and Planetary Sciences 3, 213-249.
Show in context Unambiguous identification of organic microfossils in the early geological record has always been challenging (Schopf, 1975; Knoll, 2015).
View in article
Schopf, J.W., Packer, B.M. (1987) Early Archean (3.3-billion to 3.5-billion-year-old) microfossils from Warrawoona Group, Australia. Science 237, 70-73.
Show in context There, the disputed microstructures have been interpreted as either remnants of Earth’s oldest cellular life (Schopf and Packer, 1987; Ueno et al., 2001; Schopf et al., 2002, 2018) or as abiotic organo-mineral structures (Brasier et al., 2002; Marshall et al., 2011; Wacey et al., 2016).
View in article
Schopf, J.W., Kudryavtsev, A.B., Agresti, D.G., Wdowiak, T.J., Czaja, A.D. (2002) Laser–Raman imagery of Earth's earliest fossils. Nature 416, 73-76.
Show in context There, the disputed microstructures have been interpreted as either remnants of Earth’s oldest cellular life (Schopf and Packer, 1987; Ueno et al., 2001; Schopf et al., 2002, 2018) or as abiotic organo-mineral structures (Brasier et al., 2002; Marshall et al., 2011; Wacey et al., 2016).
View in article
Schopf, J.W., Kitajima, K., Spicuzza, M.J., Kudryavtsev, A.B., Valley, J.W. (2018) SIMS analyses of the oldest known assemblage of microfossils document their taxon-correlated carbon isotope compositions. Proceedings of the National Academy of Sciences 115, 53-58.
Show in context There, the disputed microstructures have been interpreted as either remnants of Earth’s oldest cellular life (Schopf and Packer, 1987; Ueno et al., 2001; Schopf et al., 2002, 2018) or as abiotic organo-mineral structures (Brasier et al., 2002; Marshall et al., 2011; Wacey et al., 2016).
View in article
Isotopic investigation aside (Ueno et al., 2001; Schopf et al., 2018), molecular appraisal of the biogenicity of the Apex microstructures has proven inconclusive, hindered because the analyses were performed on bulk powdered samples rather than targeting the microstructures themselves (De Gregorio et al., 2009).
View in article
Shapiro, R., Konhauser, K. (2015) Hematite‐coated microfossils: primary ecological fingerprint or taphonomic oddity of the Paleoproterozoic? Geobiology 13, 209-224.
Show in context This was the case for the Strelley Pool chert, as supported by the absence of secondary minerals, but not for the most hydrothermally altered Gunflint cherts, as evidenced by the presence of iron oxides that likely formed during diagenesis (Shapiro and Konhauser, 2015; Alleon et al., 2016a).
View in article
Sugitani, K., Mimura, K., Nagaoka, T., Lepot, K., Takeuchi, M. (2013) Microfossil assemblage from the 3400Ma Strelley Pool Formation in the Pilbara Craton, Western Australia: Results form a new locality. Precambrian Research 226, 59-74.
Show in context These cherts are thought to have formed through the crystallisation of colloidal silica that was deposited in a shallow water marine environment, with some hydrothermal inputs (Sugitani et al., 2013).
View in article
Since their formation, these cherts have only undergone low grade metamorphism (prehnite-pumpellyite to lower greenschist facies; Lepot et al., 2013; Sugitani et al., 2013).
View in article
The Strelley Pool microfossils display morphologically diverse microstructures (Wacey et al., 2011a; Sugitani et al., 2013), featuring 13C-depleted carbon (Lepot et al., 2013), and are sometimes associated with pyrites that exhibit sulphur isotope signatures consistent with a biological origin (Wacey et al., 2011b).
View in article
Consistent with previous observations of thin sections (Lepot et al., 2013; Sugitani et al., 2013; Fig. 1b, h), SEM observations reveal the presence of lenticular and film-like organic microfossils present on freshly fractured faces of the PAN1-1A chert sample (Fig. 1d–g, i).
View in article
Application of the RSCM (Raman Spectroscopy of Carbonaceous Material) geothermometer (Beyssac et al., 2002; Lahfid et al., 2010) indicates that the Strelley Pool microfossils experienced peak temperatures of approximately 300 °C, which is in good agreement with regional metamorphic histories (Wacey et al., 2011a; Lepot et al., 2013; Sugitani et al., 2013).
View in article
Figure 1 [...](a) SEM image of a lenticular Strelley Pool microfossil, (b) optical photomicrograph of Strelley Pool lenticular microfossils (Sugitani et al., 2013) and (c) model morphological reconstruction in clay.
View in article
Figure 1 [...] (d–g) SEM images of Strelley Pool film-like microfossils, (h) optical photomicrograph of a film-like Strelley Pool microfossil (Sugitani et al., 2013) and (i) an enlargement of the film-like microfossil shown in f that looks like a tubular structure.
View in article
Altogether, consistent with previous claims (Lepot et al., 2013; Sugitani et al., 2013), the spatially resolved chemical information reported here supports the biogenicity of the Strelley Pool microfossils, advocating them as the oldest currently known authentic microfossils.
View in article
Ueno, Y., Isozaki, Y., Yurimoto, H., Maruyama, S. (2001) Carbon isotopic signatures of individual Archean microfossils (?) from Western Australia. International Geology Review 43, 196-212.
Show in context There, the disputed microstructures have been interpreted as either remnants of Earth’s oldest cellular life (Schopf and Packer, 1987; Ueno et al., 2001; Schopf et al., 2002, 2018) or as abiotic organo-mineral structures (Brasier et al., 2002; Marshall et al., 2011; Wacey et al., 2016).
View in article
Isotopic investigation aside (Ueno et al., 2001; Schopf et al., 2018), molecular appraisal of the biogenicity of the Apex microstructures has proven inconclusive, hindered because the analyses were performed on bulk powdered samples rather than targeting the microstructures themselves (De Gregorio et al., 2009).
View in article
Wacey, D., Kilburn, M.R., Saunders, M., Cliff, J., Brasier, M.D. (2011a) Microfossils of sulphur-metabolizing cells in 3.4-billion-year-old rocks of Western Australia. Nature Geoscience 4, 698-702.
Show in context The Strelley Pool microfossils display morphologically diverse microstructures (Wacey et al., 2011a; Sugitani et al., 2013), featuring 13C-depleted carbon (Lepot et al., 2013), and are sometimes associated with pyrites that exhibit sulphur isotope signatures consistent with a biological origin (Wacey et al., 2011b).
View in article
Application of the RSCM (Raman Spectroscopy of Carbonaceous Material) geothermometer (Beyssac et al., 2002; Lahfid et al., 2010) indicates that the Strelley Pool microfossils experienced peak temperatures of approximately 300 °C, which is in good agreement with regional metamorphic histories (Wacey et al., 2011a; Lepot et al., 2013; Sugitani et al., 2013).
View in article
Wacey, D., Saunders, M., Brasier, M.D., Kilburn, M.R. (2011b) Earliest microbially mediated pyrite oxidation in ~3.4 billion-year-old sediments. Earth and Planetary Science Letters 301, 393-402.
Show in context The Strelley Pool microfossils display morphologically diverse microstructures (Wacey et al., 2011a; Sugitani et al., 2013), featuring 13C-depleted carbon (Lepot et al., 2013), and are sometimes associated with pyrites that exhibit sulphur isotope signatures consistent with a biological origin (Wacey et al., 2011b).
View in article
Wacey, D., Saunders, M., Kong, C., Brasier, A., Brasier, M. (2016) 3.46 Ga Apex chert ‘microfossils’ reinterpreted as mineral artefacts produced during phyllosilicate exfoliation. Gondwana Research 36, 296-313.
Show in context There, the disputed microstructures have been interpreted as either remnants of Earth’s oldest cellular life (Schopf and Packer, 1987; Ueno et al., 2001; Schopf et al., 2002, 2018) or as abiotic organo-mineral structures (Brasier et al., 2002; Marshall et al., 2011; Wacey et al., 2016).
View in article
top
Supplementary Information
The Supplementary Information includes:
- Methods
- Supplementary Information References
Download the Supplementary Information (PDF).
Figures and Tables
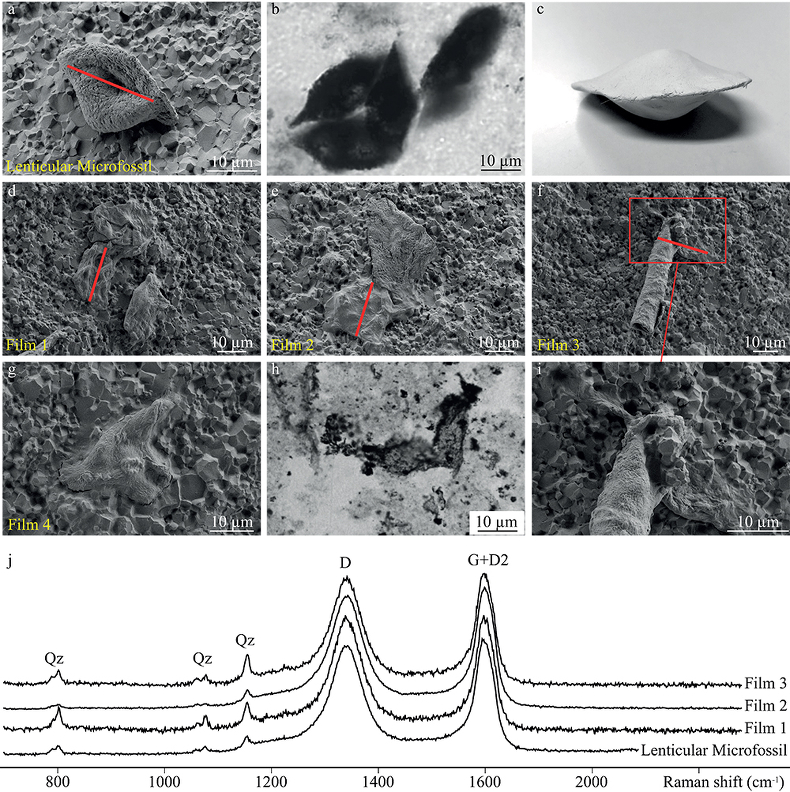
Figure 1 Optical photomicrographs, SEM images and Raman spectra of the 3.4 Ga Strelley Pool organic microfossils observed on a freshly fractured chert fragment. (a) SEM image of a lenticular Strelley Pool microfossil, (b) optical photomicrograph of Strelley Pool lenticular microfossils (Sugitani et al., 2013
Sugitani, K., Mimura, K., Nagaoka, T., Lepot, K., Takeuchi, M. (2013) Microfossil assemblage from the 3400Ma Strelley Pool Formation in the Pilbara Craton, Western Australia: Results form a new locality. Precambrian Research 226, 59-74.
) and (c) model morphological reconstruction in clay. (d–g) SEM images of Strelley Pool film-like microfossils, (h) optical photomicrograph of a film-like Strelley Pool microfossil (Sugitani et al., 2013Sugitani, K., Mimura, K., Nagaoka, T., Lepot, K., Takeuchi, M. (2013) Microfossil assemblage from the 3400Ma Strelley Pool Formation in the Pilbara Craton, Western Australia: Results form a new locality. Precambrian Research 226, 59-74.
) and (i) an enlargement of the film-like microfossil shown in f that looks like a tubular structure. Red lines indicate where the FIB sections were extracted for TEM and STXM analyses (Film 1–4). (j) Raman spectra of the Strelley Pool organic microfossils These spectra are typical of disordered carbon having experienced 300 °C, with the D and composite G + D2 bands displaying similar intensities. Qz corresponds to quartz.
Figure 2 TEM analyses of the Strelley Pool organic microfossils. STEM image of the lenticular microfossil (a) shown in Figure 1a and of the two film-like microfossils (b–c) shown in Figure 1d and 1f, respectively. Organic carbon (which appears darker than mineral phases) is closely associated with micro-quartz, clays and crystals of chlorides and sulphates. The main chemical elements are indicated. (d) TEM-EDX spectra of the organic microfossils and the associated mineral phases.
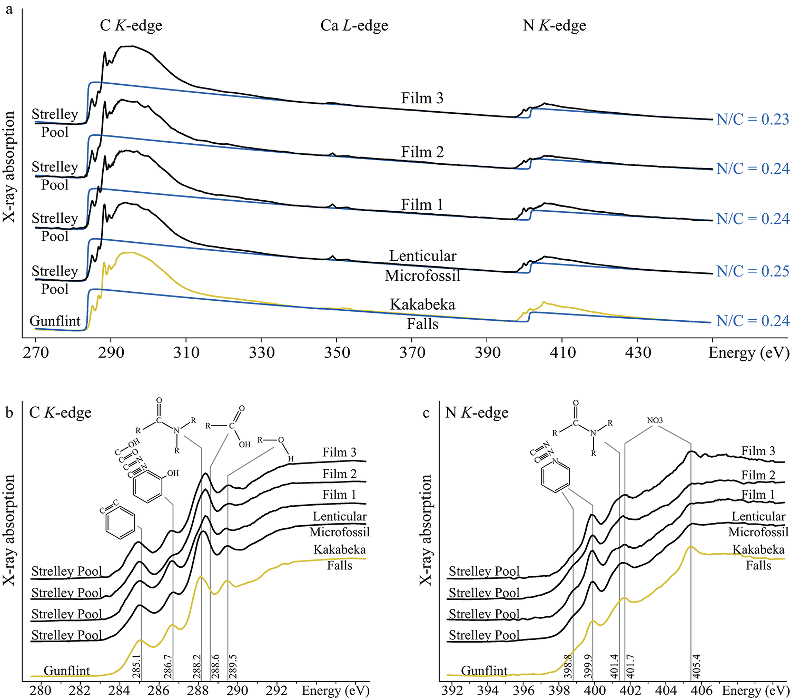
Figure 3 X-ray absorption analyses of the Strelley Pool organic microfossils shown in Figures 1 and 2 and corresponding N/C values. (a) X-ray absorption near edge structure (XANES) spectra at the carbon K-edge and (b) nitrogen K-edge. (c) Diagnostic absorption features include: 285.1 eV for aromatic/olefinic groups, 286.7 eV for imine/nitrile/carbonyl/phenol groups, 288.2 eV for amide groups, 288.6 eV for carboxyl/ester/acetal groups, 289.4 eV for hydroxyl groups, 398.8–399.9 eV - imine/nitrile/aromatic groups, 401.4 eV for amide groups, 401.7 and 405.4 eV for potassium and/or calcium nitrates. A spectrum of a 1.9 Ga Gunflint microfossil (Alleon et al., 2016a
Alleon, J., Bernard, S., Le Guillou, C., Marin-Carbonne, J., Pont, S., Beyssac, O., McKeegan, K.D., Robert, F. (2016a) Molecular preservation of 1.88 Ga Gunflint organic microfossils as a function of temperature and mineralogy. Nature Communications 7, 11977.
) is shown for comparison.