



Siderophores and the formation of cerium anomalies in anoxic environments
Affiliations | Corresponding Author | Cite as | Funding informationPublished by the European Association of Geochemistry
under Creative Commons License CC BY-NC-ND 4.0
Keywords: Ce anomalies, siderophores, Ce oxidation, rare earth elements, geochemical proxies, redox proxies, early Earth

- Share this article




-
Article views:125Cumulative count of HTML views and PDF downloads.
- Download Citation
- Rights & Permissions

top
Abstract
Figures
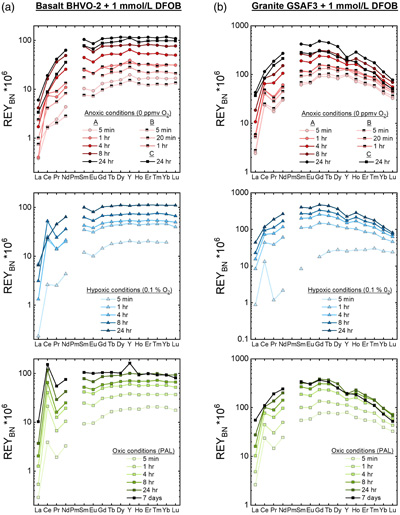 Figure 1 REYBN patterns of time series experiments with (a) basalt and (b) granite. | 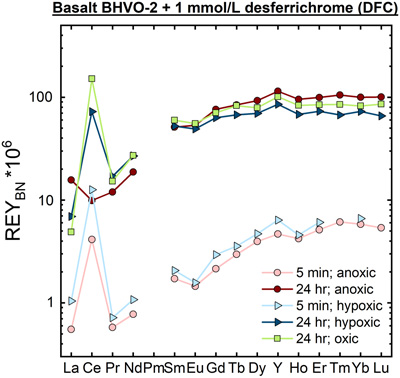 Figure 2 REYBN patterns of experiments with basalt and DFC. | 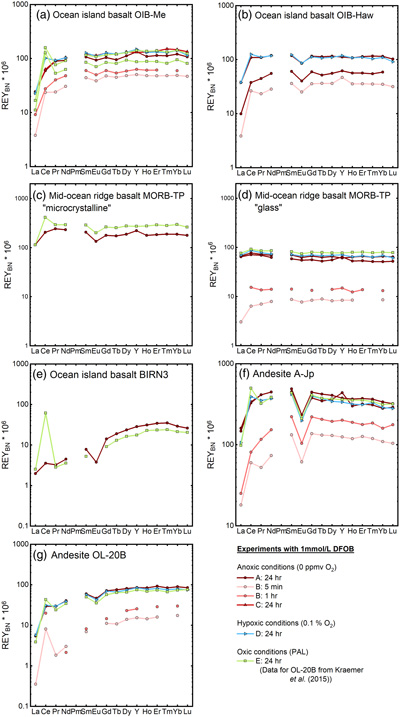 Figure 3 REYBN patterns of leaching experiments with different igneous rocks. The letters A-E in the legend refer to different experiment runs. Details are listed in Table S-2. | 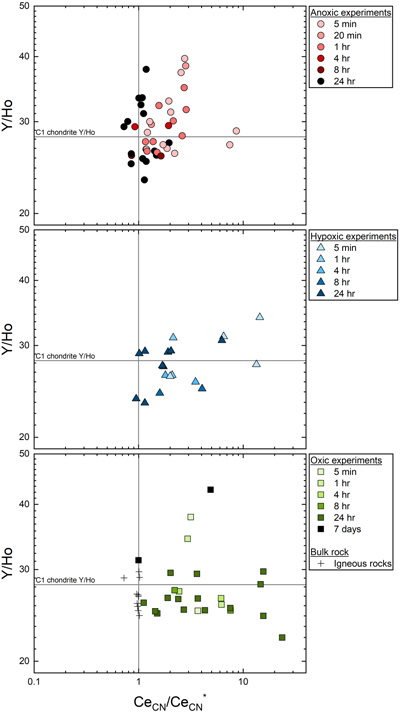 Figure 4 Graph of CeCN/CeCN* vs. Y/Ho ratio for bulk rocks and leachates. |
| Figure 1 | Figure 2 | Figure 3 | Figure 4 |
top
Introduction
Cerium belongs to the lanthanoides, which, together with the pseudolanthanoid Y, are referred to as “rare earth elements and yttrium” (REY). The REY show coherent behaviour in the natural environment due to their similar charge and systematically decreasing ionic radii. Cerium is an exception, as it can be oxidised to Ce(IV) and hence decoupled from its redox-insensitive, strictly trivalent REY neighbors La and Pr. During pedogenesis and oxidative weathering, Ce(IV) may form insoluble cerianite (CeO2) which remains fixed in the regolith when the REY(III) are gradually mobilised (Braun et al., 1990
Braun, J.-J., Pagel, M., Muller, J.-P., Bilong, P., Michard, A., Guillet, B. (1990) Cerium anomalies in lateritic profiles. Geochimica et Cosmochimica Acta 54, 781–795. https://doi.org/10.1016/0016-7037(90)90373-S
), eventually producing Ce anomalies in normalised REY patterns of soil. Likewise, Fe and Mn-oxyhydroxides have the tendency to surface-oxidise Ce upon sorption, facilitating positive CeN anomalies at the mineral surfaces and, with time, progressively more negative anomalies in the associated (pore) solutions (Bau and Koschinsky, 2009Bau, M., Koschinsky, A. (2009) Oxidative scavenging of cerium on hydrous Fe oxide: Evidence from the distribution of rare earth elements and yttrium between Fe oxides and Mn oxides in hydrogenetic ferromanganese crusts. Geochemical Journal 43, 37–47. https://doi.org/10.2343/geochemj.1.0005
). Due to their particle reactivity and the presumed immobility of REY, presence or absence of Ce anomalies in palaeoweathering profiles and marine chemical sediments are commonly used as a robust qualitative redox proxy (e.g., German and Elderfield, 1990German, C.R., Elderfield, H. (1990) Application of the Ce anomaly as a paleoredox indicator: The ground rules. Paleoceanography and Paleoclimatology 5, 823–833. https://doi.org/10.1029/PA005i005p00823
; Riding et al., 2014Riding, R., Fralick, P., Liang, L. (2014) Identification of an Archean marine oxygen oasis. Precambrian Research 251, 232–237. https://doi.org/10.1016/j.precamres.2014.06.017
).Biota facilitates a strong control on redox-sensitive elements, such as Fe (e.g., Byrne et al., 2015
Byrne, J.M., Klueglein, N., Pearce, C., Rosso, K.M., Appel, E., Kappler, A. (2015) Redox cycling of Fe(II) and Fe(III) in magnetite by Fe-metabolizing bacteria. Science 347, 1473–1476. https://doi.org/10.1126/science.aaa4834
). A class of biogenic ligands that interacts strongly with oxide and silicate minerals (Brantley et al., 2001Brantley, S.L., Liermann, L., Bau, M., Wu, S. (2001) Uptake of Trace Metals and Rare Earth Elements from Hornblende by a Soil Bacterium. Geomicrobiology Journal 18, 37–61. https://doi.org/10.1080/01490450151079770
; Kraemer et al., 2014Kraemer, S.M., Duckworth, O.W., Harrington, J.M., Schenkeveld, W.D.C. (2014) Metallophores and Trace Metal Biogeochemistry. Aquatic Geochemistry 21, 159–195. https://doi.org/10.1007/s10498-014-9246-7
) and that solubilises Fe, REY and other high field strength elements (Yoshida et al., 2004Yoshida, T., Ozaki, T., Ohnuki, T., Francis, A.J. (2004) Adsorption of rare earth elements by γ-Al2O3 and Pseudomonas fluorescens cells in the presence of desferrioxamine B: implication of siderophores for the Ce anomaly. Chemical Geology 212, 239–246. https://doi.org/10.1016/j.chemgeo.2004.08.046
; Christenson and Schijf, 2011Christenson, E.A., Schijf, J. (2011) Stability of YREE complexes with the trihydroxamate siderophore desferrioxamine B at seawater ionic strength. Geochimica et Cosmochimica Acta 75, 7047–7062. https://doi.org/10.1016/j.gca.2011.09.022
; Bau et al., 2013Bau, M., Tepe, N., Mohwinkel, D. (2013) Siderophore-promoted transfer of rare earth elements and iron from volcanic ash into glacial meltwater, river and ocean water. Earth and Planetary Science Letters 364, 30–36. https://doi.org/10.1016/j.epsl.2013.01.002
; Kraemer et al., 2015Kraemer, D., Kopf, S., Bau, M. (2015) Oxidative mobilization of cerium and uranium and enhanced release of “immobile” high field strength elements from igneous rocks in the presence of the biogenic siderophore desferrioxamine B. Geochimica et Cosmochimica Acta 165, 263–279. https://doi.org/10.1016/j.gca.2015.05.046
, 2017Kraemer, D., Tepe, N., Pourret, O., Bau, M. (2017) Negative cerium anomalies in manganese (hydr)oxide precipitates due to cerium oxidation in the presence of dissolved siderophores. Geochimica et Cosmochimica Acta 196, 197–208. https://doi.org/10.1016/j.gca.2016.09.018
) are the so-called “siderophores”. These are excreted by bacteria, plants and fungi, primarily to mobilise bioessential Fe (Haas, 2003Haas, H. (2003) Molecular genetics of fungal siderophore biosynthesis and uptake: the role of siderophores in iron uptake and storage. Applied Microbiology and Biotechnology 62, 316–330. https://doi.org/10.1007/s00253-003-1335-2
) and to cope with metal toxicity (Braud et al., 2010Braud, A., Geoffroy, V., Hoegy, F., Mislin, G.L.A., Schalk, I.J. (2010) Presence of the siderophores pyoverdine and pyochelin in the extracellular medium reduces toxic metal accumulation in Pseudomonas aeruginosa and increases bacterial metal tolerance. Environmental Microbiology Reports 2, 419–425. https://doi.org/10.1111/j.1758-2229.2009.00126.x
). Siderophore-promoted oxidation of Ce under oxic conditions has been described in literature (Brantley et al., 2001Brantley, S.L., Liermann, L., Bau, M., Wu, S. (2001) Uptake of Trace Metals and Rare Earth Elements from Hornblende by a Soil Bacterium. Geomicrobiology Journal 18, 37–61. https://doi.org/10.1080/01490450151079770
; Yoshida et al., 2004Yoshida, T., Ozaki, T., Ohnuki, T., Francis, A.J. (2004) Adsorption of rare earth elements by γ-Al2O3 and Pseudomonas fluorescens cells in the presence of desferrioxamine B: implication of siderophores for the Ce anomaly. Chemical Geology 212, 239–246. https://doi.org/10.1016/j.chemgeo.2004.08.046
; Bau et al., 2013Bau, M., Tepe, N., Mohwinkel, D. (2013) Siderophore-promoted transfer of rare earth elements and iron from volcanic ash into glacial meltwater, river and ocean water. Earth and Planetary Science Letters 364, 30–36. https://doi.org/10.1016/j.epsl.2013.01.002
; Kraemer et al., 2015Kraemer, D., Kopf, S., Bau, M. (2015) Oxidative mobilization of cerium and uranium and enhanced release of “immobile” high field strength elements from igneous rocks in the presence of the biogenic siderophore desferrioxamine B. Geochimica et Cosmochimica Acta 165, 263–279. https://doi.org/10.1016/j.gca.2015.05.046
, 2017Kraemer, D., Tepe, N., Pourret, O., Bau, M. (2017) Negative cerium anomalies in manganese (hydr)oxide precipitates due to cerium oxidation in the presence of dissolved siderophores. Geochimica et Cosmochimica Acta 196, 197–208. https://doi.org/10.1016/j.gca.2016.09.018
).This study fills the important knowledge gap on the impact of the oxygen level of a system on this process. The naturally abundant microbial siderophore desferrioxamine B (DFOB), produced by Streptomyces pilosus, and the fungal siderophore desferrichrome (DFC), produced by Ustilago sphaerogena, are used as model siderophores for batch incubation experiments with powdered igneous rocks to simulate water–rock interaction (WRI) at oxygen levels of (i) 0 ppmv O2 (‘anoxic’), (ii) 0.1 % O2 (‘hypoxic’) and (iii) ∼21 % O2 (present atmospheric level; ‘oxic’). The experiments were conducted at room temperature in a gloveless anaerobic chamber with various sampling times and a fixed rock/water ratio of 20 g/L.
top
Results
A set of igneous rocks, comprised of mid-ocean ridge basalt (MORB; glass and microcrystalline), island-arc basalt (IAB), andesite and granite, was chosen for investigating WRI at different atmospheric conditions. Concentrations of REY in leachates were normalised to those in the respective bulk rock (subscript “BN”) to facilitate comparison between mobilised REY relative to the bulk rock inventory. We emphasise that the rock samples were pristine and that the bulk rock REY patterns (Fig. S-1) did not show any signs of alteration.
Figures 1–3 show REYBN patterns of the leaching experiments at different run times, and Figure 4 shows calculated Ce anomalies (CeCN/CeCN*) vs. Y/Ho ratio of the anoxic (shades of red, circles), hypoxic (shades of blue, triangles) and oxic experiments (shades of green, squares). Leaching of REY from igneous rocks in the presence of either of the two model siderophores under the three studied pO2 conditions revealed the following:
- (i) the amount of mobilised REY increases with time;
- (ii) HREY mobilisation is stronger than LREY mobilisation, especially La is depleted;
- (iii) patterns of the anoxic to oxic experiments at the same time interval are similar, except for Ce, and corroborate published data from oxic experiments (Kraemer et al., 2015
Kraemer, D., Kopf, S., Bau, M. (2015) Oxidative mobilization of cerium and uranium and enhanced release of “immobile” high field strength elements from igneous rocks in the presence of the biogenic siderophore desferrioxamine B. Geochimica et Cosmochimica Acta 165, 263–279. https://doi.org/10.1016/j.gca.2015.05.046
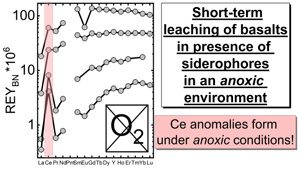
); - (iv) in the first 5 min of leaching, positive CeBN anomalies develop even under strictly anoxic conditions, regardless of mineralogical composition, texture and REY concentrations of the rock sample; after 60 min at the latest, these CeBN anomalies become smaller, and later disappear in all anoxic experiments, resulting in patterns without CeBN anomaly after 24 hr;
- (v) positive CeBN anomalies formed in all hypoxic and oxic experiments after at least 5 min, except in experiments with glassy MORB and granite. Nevertheless, the CeBN anomalies that formed during the hypoxic experiments are less pronounced than those that developed during the oxic experiments, suggesting that pO2 exerts a control on the formation of Ce anomalies during WRI with siderophores;
- (vi) Y-Ho fractionation was more pronounced in the anoxic experiments than in the other experiments (Fig. 4).
Figure 1 REYBN patterns of time series experiments with (a) basalt and (b) granite.
Figure 2 REYBN patterns of experiments with basalt and DFC.
Figure 3 REYBN patterns of leaching experiments with different igneous rocks. The letters A-E in the legend refer to different experiment runs. Details are listed in Table S-2.
Figure 4 Graph of CeCN/CeCN* vs. Y/Ho ratio for bulk rocks and leachates.
In the course of individual experiments, pH first decreased from 5.5 to <5, then increased and mostly remained in the range of 5–7 after 24 hr (Figs. S-2–S-4, Table S-4), similar to previous studies (Kraemer et al., 2015
Kraemer, D., Kopf, S., Bau, M. (2015) Oxidative mobilization of cerium and uranium and enhanced release of “immobile” high field strength elements from igneous rocks in the presence of the biogenic siderophore desferrioxamine B. Geochimica et Cosmochimica Acta 165, 263–279. https://doi.org/10.1016/j.gca.2015.05.046
). Some experiments reached pH values of 8 and higher. Cerium anomaly formation, especially under anoxic conditions, however, was not correlated with final solution pH (Figs. S-2, S-3). Leaching under anoxic and hypoxic conditions caused a generally slower pH increase over time (Fig. S-4). Over the observed pH range, DFOB speciation was dominated by H4DFOB+ species with minor to equal contributions of H3DFOB (Fig. S-5).Control experiments with DI and 0.01 mol/L HCl under anoxic conditions did not produce Ce anomalies in the leachate solutions (Fig. S-6).
top
Discussion
The experiments indicate that leaching of redox-insensitive REY(III) by both DFOB and DFC is largely independent of actual pO2 as evidenced by very similar patterns between the individual experiments with the same run times. The patterns between the different experimental sets overlap, except for redox-sensitive Ce, implying that complexation of redox-insensitive trace elements with DFOB or DFC is solely a function of the complex stability constants, the mineralogical association of the REY, the texture of the rock, and the individual solubility of the host minerals, but not of oxygen. Due to the scarcity of available data for complexation of REY with DFC, the following discussion focuses on REY-DFOB interaction. Stability constants for REY-DFOB complexes are several orders of magnitude higher than those of mono- and dicarbonate REY complexes in seawater (Liu and Byrne, 1998
Liu, X., Byrne, R.H. (1998) Comprehensive Investigation of Yttrium and Rare Earth Element Complexation by Carbonate Ions Using ICP–Mass Spectrometry. Journal of Solution Chemistry 27, 803–815. https://doi.org/10.1023/A:1022677119835
), with log Kf of hexadentate complexes ranging from 10.09 for La to 15.19 for Lu at seawater ionic strength (Christenson and Schijf, 2011Christenson, E.A., Schijf, J. (2011) Stability of YREE complexes with the trihydroxamate siderophore desferrioxamine B at seawater ionic strength. Geochimica et Cosmochimica Acta 75, 7047–7062. https://doi.org/10.1016/j.gca.2011.09.022
). In comparison, the hexadentate coordination of Fe3+ results in a stability constant of 30.6 (DFOB; Martell and Smith, 2001Martell, A., Smith, R. (2001) NIST Stability Constants of Metal Complexes Database 46, v. 6.0. National Institute of Standards and Technology, Gaithersburg, MD.
). The increasing complex stability from light to heavy REY is observed, e.g., in the leachates of the basalts and andesites, as the patterns show an increase from LREYBN towards MREYBN and HREYBN.The only leachates with different REYBN patterns are of glassy MORB (Fig. 3d; flat REYBN without fractionation) and of granite (Fig. 1b; bell-shaped REYBN). The “MREY bulge” observed in the granite experiment may be related to highly insoluble minerals present in felsic plutonic rocks. The leachates of glassy MORB show lower REY concentrations than those of the microcrystalline counterpart. As REY are incompatible with respect to most rock-forming igneous minerals, they are, in rocks with porphyritic/phaneritic textures, not incorporated into the crystal lattice but are preferentially located along grain boundaries and in the interstitial crystal spaces (Giese and Bau, 1994
Giese, U., Bau, M. (1994) Trace element accessibility in mid-ocean ridge and ocean island basalt: an experimental approach. Goldschmidt Abstracts 1994, 329–330. https://rruff.info/doclib/MinMag/Volume_58A/58A-1-329.pdf
). During WRI, this REY pool is mobilised first since no dissolution of well-crystallised minerals is necessary. Divalent Eu is compatible with respect to, and incorporated into, Ca-rich plagioclase, and hence the REY pool on grain boundaries and in interstitial spaces is depleted in Eu (negative EuCN anomaly). When this pool is mobilised during WRI, the solution inherits the EuCN anomaly. In volcanic glass, however, Eu(II) and all REY(III) are homogenously distributed, and hence no EuCN anomaly can develop (Giese and Bau, 1994Giese, U., Bau, M. (1994) Trace element accessibility in mid-ocean ridge and ocean island basalt: an experimental approach. Goldschmidt Abstracts 1994, 329–330. https://rruff.info/doclib/MinMag/Volume_58A/58A-1-329.pdf
; Bach and Irber, 1998Bach, W., Irber, W. (1998) Rare earth element mobility in the oceanic lower sheeted dyke complex: evidence from geochemical data and leaching experiments. Chemical Geology 151, 309–326. https://doi.org/10.1016/S0009-2541(98)00087-4
; Shibata et al., 2006Shibata, S.-N., Tanaka, T., Yamamoto, K. (2006) Crystal structure control of the dissolution of rare earth elements in water-mineral interactions. Geochemical Journal 40, 437–446. https://doi.org/10.2343/geochemj.40.437
; Kraemer et al., 2015Kraemer, D., Kopf, S., Bau, M. (2015) Oxidative mobilization of cerium and uranium and enhanced release of “immobile” high field strength elements from igneous rocks in the presence of the biogenic siderophore desferrioxamine B. Geochimica et Cosmochimica Acta 165, 263–279. https://doi.org/10.1016/j.gca.2015.05.046
).We conclude that pO2 neither influences the amount nor the fractionation of individual or total REY(III) mobilised from igneous rocks during WRI in the presence of DFOB or DFC. Our experiments demonstrate, in accordance with published data, that the solubilisation and complexation of isovalent REY with DFOB or DFC is oxygen independent.
Leaching under hypoxic and oxic conditions leads to only minor fractionation of the Y-Ho geochemical twins, but pronounced fractionation was observed during short-term WRI under anoxic conditions (Fig. 4). The stability constants of Y and Ho with DFOB are similar (Christenson and Schijf, 2011
Christenson, E.A., Schijf, J. (2011) Stability of YREE complexes with the trihydroxamate siderophore desferrioxamine B at seawater ionic strength. Geochimica et Cosmochimica Acta 75, 7047–7062. https://doi.org/10.1016/j.gca.2011.09.022
) and no significant fractionation would be expected, in contrast to our results. Yttrium, however, appears to have a higher initial accessibility during WRI than Ho (Bau et al., 1998Bau, M., Usui, A., Pracejus, B., Mita, N., Kanai, Y., Irber, W., Dulski, P. (1998) Geochemistry of low-temperature water-rock interaction: Evidence from natural waters, andesite, and iron-oxyhydroxide precipitates at Nishiki-numa iron-spring, Hokkaido, Japan. Chemical Geology 151, 293–307. https://doi.org/10.1016/S0009-2541(98)00086-2
), which could explain the Y fractionation during short-term WRI.Cerium oxidation by the siderophore DFOB under oxic conditions has been demonstrated in several studies. Yoshida et al. (2004)
Yoshida, T., Ozaki, T., Ohnuki, T., Francis, A.J. (2004) Adsorption of rare earth elements by γ-Al2O3 and Pseudomonas fluorescens cells in the presence of desferrioxamine B: implication of siderophores for the Ce anomaly. Chemical Geology 212, 239–246. https://doi.org/10.1016/j.chemgeo.2004.08.046
conducted adsorption experiments of REY-DFOB complexes on γ-Al2O3 and P. fluorescens and found that Ce behaves markedly different from La(III) and Pr(III). They demonstrated the presence of Ce(IV)-DFOB in solution and concluded an oxidation of Ce(III) to Ce(IV) during DFOB complexation/adsorption. Siderophore-mediated oxidation was also demonstrated during oxic WRI with igneous rocks (Bau et al., 2013Bau, M., Tepe, N., Mohwinkel, D. (2013) Siderophore-promoted transfer of rare earth elements and iron from volcanic ash into glacial meltwater, river and ocean water. Earth and Planetary Science Letters 364, 30–36. https://doi.org/10.1016/j.epsl.2013.01.002
; Kraemer et al., 2015Kraemer, D., Kopf, S., Bau, M. (2015) Oxidative mobilization of cerium and uranium and enhanced release of “immobile” high field strength elements from igneous rocks in the presence of the biogenic siderophore desferrioxamine B. Geochimica et Cosmochimica Acta 165, 263–279. https://doi.org/10.1016/j.gca.2015.05.046
) and during scavenging onto Mn (hydr)oxides (Kraemer et al., 2017Kraemer, D., Tepe, N., Pourret, O., Bau, M. (2017) Negative cerium anomalies in manganese (hydr)oxide precipitates due to cerium oxidation in the presence of dissolved siderophores. Geochimica et Cosmochimica Acta 196, 197–208. https://doi.org/10.1016/j.gca.2016.09.018
) with the development of a positive CeN anomaly.In contrast to the log Kf of REY(III)-DFOB (between 10.09 ± 0.08 and 15.19 ± 0.02; Christenson and Schijf, 2011
Christenson, E.A., Schijf, J. (2011) Stability of YREE complexes with the trihydroxamate siderophore desferrioxamine B at seawater ionic strength. Geochimica et Cosmochimica Acta 75, 7047–7062. https://doi.org/10.1016/j.gca.2011.09.022
) and Ce(III)-DFOB hexadentate complexes (log Kf ≈ 11.59; back-extrapolated from Nd(III)- and Pr(III)-DFOB data of Christenson and Schijf, 2011Christenson, E.A., Schijf, J. (2011) Stability of YREE complexes with the trihydroxamate siderophore desferrioxamine B at seawater ionic strength. Geochimica et Cosmochimica Acta 75, 7047–7062. https://doi.org/10.1016/j.gca.2011.09.022
), log Kf of Ce(IV)-DFOB was estimated to be significantly higher (25–30; Yoshida et al., 2004Yoshida, T., Ozaki, T., Ohnuki, T., Francis, A.J. (2004) Adsorption of rare earth elements by γ-Al2O3 and Pseudomonas fluorescens cells in the presence of desferrioxamine B: implication of siderophores for the Ce anomaly. Chemical Geology 212, 239–246. https://doi.org/10.1016/j.chemgeo.2004.08.046
) and thus comparable to Fe(III)-DFOB. The DFOB-mediated oxidation of redox-sensitive elements under oxic conditions was explained as follows:- (i) a lowering of the oxidation potential by DFOB-complexation and rapid oxidation by ambient air (“air-oxidation”; Duckworth and Sposito, 2005
Duckworth, O.W., Sposito, G. (2005) Siderophore–Manganese (III) interactions. I. Air-Oxidation of Manganese(II) Promoted by Desferrioxamine B. Environmental Science and Technology 39, 6037–6044. https://doi.org/10.1021/es050275k
) which increases the metal-ligand complex stability (Hernlem et al., 1999Hernlem, B.J., Vane, L.M., Sayles, G.D. (1999) The application of siderophores for metal recovery and waste remediation: examination of correlations for prediction of metal affinities. Water Research 33, 951–960. https://doi.org/10.1016/S0043-1354(98)00293-0
; Kraemer et al., 2014Kraemer, S.M., Duckworth, O.W., Harrington, J.M., Schenkeveld, W.D.C. (2014) Metallophores and Trace Metal Biogeochemistry. Aquatic Geochemistry 21, 159–195. https://doi.org/10.1007/s10498-014-9246-7
), and - (ii) a disequilibrium between Ce3+aq and Ce4+aq at the mineral–water interface, where minute amounts of tetravalent Ce in solution are strongly complexed by DFOB, facilitating further disequilibrium and Ce4+ regeneration by continuous oxidation of Ce3+ to maintain redox equilibrium (“siderophore redox pump”; Bau et al., 2013
Bau, M., Tepe, N., Mohwinkel, D. (2013) Siderophore-promoted transfer of rare earth elements and iron from volcanic ash into glacial meltwater, river and ocean water. Earth and Planetary Science Letters 364, 30–36. https://doi.org/10.1016/j.epsl.2013.01.002
; Kraemer et al., 2015Kraemer, D., Kopf, S., Bau, M. (2015) Oxidative mobilization of cerium and uranium and enhanced release of “immobile” high field strength elements from igneous rocks in the presence of the biogenic siderophore desferrioxamine B. Geochimica et Cosmochimica Acta 165, 263–279. https://doi.org/10.1016/j.gca.2015.05.046
, 2017Kraemer, D., Tepe, N., Pourret, O., Bau, M. (2017) Negative cerium anomalies in manganese (hydr)oxide precipitates due to cerium oxidation in the presence of dissolved siderophores. Geochimica et Cosmochimica Acta 196, 197–208. https://doi.org/10.1016/j.gca.2016.09.018
).
Here, a Ce-LREY fractionation was observed even under anoxic conditions, although confined to short-term WRI. Clearly, the mechanisms proposed for Ce oxidation under (hyp)oxic conditions fail to explain this observation.
At the observed pH range, H4DFOB+ is the dominant species in solution, and anoxic Ce oxidation may follow a reaction path similar to the anoxic siderophore-promoted oxidation of ferrous Fe (Farkas et al., 2001
Farkas, E., Enyedy, E.A., Zékány, L., Deák, G. (2001) Interaction between iron(II) and hydroxamic acids: oxidation of iron(II) to iron(III) by desferrioxamine B under anaerobic conditions. Journal of Inorganic Biochemistry 83, 107–114. https://doi.org/10.1016/S0162-0134(00)00197-5
). The proposed reaction may be rewritten for a postulated anoxic Ce3+ oxidation process: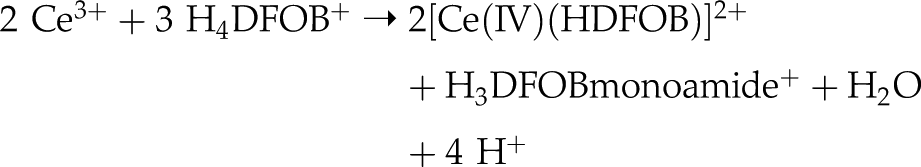
The results show that Ce fractionation under anoxic conditions is limited to short-term WRI. Apparently, only “free” DFOB molecules may participate in this redox reaction. Hence, this process is limited to the first minutes of leaching, when the activity of “free” H4DFOB+ is high and complexation with other metals with high complex stability (in particular Fe, which is present in excessive amounts in igneous rocks and which was also mobilised during the experiments; Figs. S-7–S-9, Table S-4) had not yet reduced the initial availability of this ligand. As WRI proceeds, the minute amounts of Ce(IV)-DFOB complexes formed under anoxic conditions are concealed by continuously increasing amounts of LREY(III)-DFOB complexes (including Ce(III)-DFOB) which effectively hide the Ce(IV) compounds in the REYBN patterns.
We showed experimentally that complexation by biogenic ligands such as DFOB/DFC provides an anoxic and hypoxic pathway for the oxidation of Ce3+, the subsequent oxidative decoupling of Ce from its neighbours in the REY series, and eventually the formation of positive Ce anomalies in normalised REY patterns of solutions produced during low-temperature WRI. We emphasise, however, that features of the Ce anomalies (e.g., height of peak or relative fractionation from Pr(III) or Nd(III)), cannot be used to distinguish between redox-driven and siderophore-driven fractionation.
In modern seawater, complexation of bioessential micronutrients (e.g., Fe(III)) is dominated by organic ligands, and previous studies have suggested that this ligand pool is dominated by siderophores (van den Berg, 1995
van den Berg, C.M.G. (1995) Evidence for organic complexation of iron in seawater. Marine Chemistry 50, 139–157. https://doi.org/10.1016/0304-4203(95)00032-M
; Gledhill et al., 1998Gledhill, M., van den Berg, C.M.G., Nolting, R.F., Timmermans, K.R. (1998) Variability in the speciation of iron in the northern North Sea. Marine Chemistry 59, 283–300. https://doi.org/10.1016/S0304-4203(97)00097-2
; Mawji et al., 2008Mawji, E., Gledhill, M., Milton, J.A., Tarran, G.A., Ussher, S., Thompson, A., Wolff, G.A., Worsfold, P.J., Achterberg, E.P. (2008) Hydroxamate Siderophores: Occurrence and Importance in the Atlantic Ocean. Environmental Science & Technology 42, 8675–8680. https://doi.org/10.1021/es801884r
). Recent literature suggests that cyanobacteria may have produced siderophores for Fe(III) acquisition even before the Great Oxidation Event in the Palaeoproterozoic (Enzingmüller-Bleyl et al., 2021Enzingmüller-Bleyl, T.C., Boden, J.S., Herrmann, A.J., Ebel, K.W., Sánchez-Baracaldo, P., Frankenberg-Dinkel, N., Gehringer, M.M. (2021) Lack of Fe(II) transporters in basal Cyanobacteria complicates iron uptake in ferruginous Archean oceans. (Preprint) bioRxiv, 2021.11.08.467730. https://doi.org/10.1101/2021.11.08.467730
). Today, siderophores also enable microorganisms to reduce toxic metal accumulation and to increase metal tolerance (e.g., Braud et al., 2010Braud, A., Geoffroy, V., Hoegy, F., Mislin, G.L.A., Schalk, I.J. (2010) Presence of the siderophores pyoverdine and pyochelin in the extracellular medium reduces toxic metal accumulation in Pseudomonas aeruginosa and increases bacterial metal tolerance. Environmental Microbiology Reports 2, 419–425. https://doi.org/10.1111/j.1758-2229.2009.00126.x
), which might have been evolutionary advantageous also for early life. Neaman et al. (2005)Neaman, A., Chorover, J., Brantley, S.L. (2005) Implications of the evolution of organic acid moieties for basalt weathering over geological time. American Journal of Science 305, 147–185. https://doi.org/10.2475/ajs.305.2.147
argued that organisms have had to cope with low nutrient bioavailability since at least the colonisation of the land surface. In (modern) soils, siderophores are at least as abundant as in marine systems, with concentrations up to the mmol/L range (Nelson et al., 1988Nelson, M., Cooper, C.R., Crowley, D.E., Reid, C.P.P., Szaniszlo, P.J. (1988) An escherichia coli bioassay of individual siderophores in soil. Journal of Plant Nutrition 11, 915–924. https://doi.org/10.1080/01904168809363856
), and significantly enhance the mobility of trace elements such as REY (Wiche et al., 2017Wiche, O., Tischler, D., Fauser, C., Lodemann, J., Heilmeier, H. (2017) Effects of citric acid and the siderophore desferrioxamine B (DFO-B) on the mobility of germanium and rare earth elements in soil and uptake in Phalaris arundinacea. International Journal of Phytoremediation 19, 746–754. https://doi.org/10.1080/15226514.2017.1284752
). More than 500 different natural siderophore compounds have been described from natural systems (Boukhalfa and Crumbliss, 2002Boukhalfa, H., Crumbliss, A.L. (2002) Chemical aspects of siderophore mediated iron transport. Biometals 15, 325–339. https://doi.org/10.1023/A:1020218608266
); yet to date, it is not possible to estimate abundances of organic ligands during early periods of Earth’s history. In this regard, biosignatures may represent an important, yet unprecise tool.As the different rock types showed very different REY fractionation patterns in the leachates, it is difficult to constrain REY patterns that are typical for WRI in presence of siderophores. The only distinct feature, apart from the Ce anomaly, is the occasionally very strong depletion of La (Bau et al., 2013
Bau, M., Tepe, N., Mohwinkel, D. (2013) Siderophore-promoted transfer of rare earth elements and iron from volcanic ash into glacial meltwater, river and ocean water. Earth and Planetary Science Letters 364, 30–36. https://doi.org/10.1016/j.epsl.2013.01.002
; Kraemer et al., 2015Kraemer, D., Kopf, S., Bau, M. (2015) Oxidative mobilization of cerium and uranium and enhanced release of “immobile” high field strength elements from igneous rocks in the presence of the biogenic siderophore desferrioxamine B. Geochimica et Cosmochimica Acta 165, 263–279. https://doi.org/10.1016/j.gca.2015.05.046
, 2017Kraemer, D., Tepe, N., Pourret, O., Bau, M. (2017) Negative cerium anomalies in manganese (hydr)oxide precipitates due to cerium oxidation in the presence of dissolved siderophores. Geochimica et Cosmochimica Acta 196, 197–208. https://doi.org/10.1016/j.gca.2016.09.018
; this study). Wang et al. (2020)Wang, X., Barrat, J.-A., Bayon, G., Chauvaud, L., Feng, D. (2020) Lanthanum anomalies as fingerprints of methanotrophy. Geochemical Perspectives Letters 14, 26–30. https://doi.org/10.7185/geochemlet.2019
identified La anomalies as fingerprints for methanotrophic activity. Therefore, we hypothesise that the observed La fractionation may also represent a characteristic biosignature of the involvement of siderophores in geochemical processes during WRI.The fractionation of Ce in the presence of biomolecules as reported here sheds new light on the application of Ce anomalies as a geochemical proxy. Besides its well-established application as a qualitative redox proxy, Ce anomalies may have the potential to be used as a “bio-proxy”, indicating the presence of (siderophore-producing) biota, potentially even under anoxic conditions.
While oxidation by bio-ligands will further complicate the use of geochemical redox proxies, this opens an exciting venue for future studies and potential applications, particularly in the field of astrobiology.
top
Acknowledgements
The very supportive editorial handling of Satish Myneni and the constructive comments of three anonymous reviewers are greatly appreciated. We acknowledge the financial support of the German Science Foundation (DFG) through grant KR4824/1-1 to D. Kraemer.
Editor: Satish Myneni
top
References
Bach, W., Irber, W. (1998) Rare earth element mobility in the oceanic lower sheeted dyke complex: evidence from geochemical data and leaching experiments. Chemical Geology 151, 309–326. https://doi.org/10.1016/S0009-2541(98)00087-4
 Show in context
Show in context In volcanic glass, however, Eu(II) and all REY(III) are homogenously distributed, and hence no EuCN anomaly can develop (Giese and Bau, 1994; Bach and Irber, 1998; Shibata et al., 2006; Kraemer et al., 2015).
View in article
Bau, M., Koschinsky, A. (2009) Oxidative scavenging of cerium on hydrous Fe oxide: Evidence from the distribution of rare earth elements and yttrium between Fe oxides and Mn oxides in hydrogenetic ferromanganese crusts. Geochemical Journal 43, 37–47. https://doi.org/10.2343/geochemj.1.0005
Show in context Likewise, Fe and Mn-oxyhydroxides have the tendency to surface-oxidise Ce upon sorption, facilitating positive CeN anomalies at the mineral surfaces and, with time, progressively more negative anomalies in the associated (pore) solutions (Bau and Koschinsky, 2009).
View in article
Bau, M., Usui, A., Pracejus, B., Mita, N., Kanai, Y., Irber, W., Dulski, P. (1998) Geochemistry of low-temperature water-rock interaction: Evidence from natural waters, andesite, and iron-oxyhydroxide precipitates at Nishiki-numa iron-spring, Hokkaido, Japan. Chemical Geology 151, 293–307. https://doi.org/10.1016/S0009-2541(98)00086-2
Show in context The stability constants of Y and Ho with DFOB are similar (Christenson and Schijf, 2011) and no significant fractionation would be expected, in contrast to our results. Yttrium, however, appears to have a higher initial accessibility during WRI than Ho (Bau et al., 1998), which could explain the Y fractionation during short-term WRI.
View in article
Bau, M., Tepe, N., Mohwinkel, D. (2013) Siderophore-promoted transfer of rare earth elements and iron from volcanic ash into glacial meltwater, river and ocean water. Earth and Planetary Science Letters 364, 30–36. https://doi.org/10.1016/j.epsl.2013.01.002
Show in context A class of biogenic ligands that interacts strongly with oxide and silicate minerals (Brantley et al., 2001; Kraemer et al., 2014) and that solubilises Fe, REY and other high field strength elements (Yoshida et al., 2004; Christenson and Schijf, 2011; Bau et al., 2013; Kraemer et al., 2015, 2017) are the so-called “siderophores”.
View in article
Siderophore-promoted oxidation of Ce under oxic conditions has been described in literature (Brantley et al., 2001; Yoshida et al., 2004; Bau et al., 2013; Kraemer et al., 2015, 2017).
View in article
Siderophore-mediated oxidation was also demonstrated during oxic WRI with igneous rocks (Bau et al., 2013; Kraemer et al., 2015) and during scavenging onto Mn (hydr)oxides (Kraemer et al., 2017) with the development of a positive CeN anomaly.
View in article
a disequilibrium between Ce3+aq and Ce4+aq at the mineral–water interface, where minute amounts of tetravalent Ce in solution are strongly complexed by DFOB, facilitating further disequilibrium and Ce4+ regeneration by continuous oxidation of Ce3+ to maintain redox equilibrium (“siderophore redox pump”; Bau et al., 2013; Kraemer et al., 2015, 2017).
View in article
The only distinct feature, apart from the Ce anomaly, is the occasionally very strong depletion of La (Bau et al., 2013; Kraemer et al., 2015, 2017; this study).
View in article
Boukhalfa, H., Crumbliss, A.L. (2002) Chemical aspects of siderophore mediated iron transport. Biometals 15, 325–339. https://doi.org/10.1023/A:1020218608266.
Show in context More than 500 different natural siderophore compounds have been described from natural systems (Boukhalfa and Crumbliss, 2002); yet to date, it is not possible to estimate abundances of organic ligands during early periods of Earth’s history.
View in article
Brantley, S.L., Liermann, L., Bau, M., Wu, S. (2001) Uptake of Trace Metals and Rare Earth Elements from Hornblende by a Soil Bacterium. Geomicrobiology Journal 18, 37–61. https://doi.org/10.1080/01490450151079770
Show in context A class of biogenic ligands that interacts strongly with oxide and silicate minerals (Brantley et al., 2001; Kraemer et al., 2014) and that solubilises Fe, REY and other high field strength elements (Yoshida et al., 2004; Christenson and Schijf, 2011; Bau et al., 2013; Kraemer et al., 2015, 2017) are the so-called “siderophores”.
View in article
Siderophore-promoted oxidation of Ce under oxic conditions has been described in literature (Brantley et al., 2001; Yoshida et al., 2004; Bau et al., 2013; Kraemer et al., 2015, 2017).
View in article
Braud, A., Geoffroy, V., Hoegy, F., Mislin, G.L.A., Schalk, I.J. (2010) Presence of the siderophores pyoverdine and pyochelin in the extracellular medium reduces toxic metal accumulation in Pseudomonas aeruginosa and increases bacterial metal tolerance. Environmental Microbiology Reports 2, 419–425. https://doi.org/10.1111/j.1758-2229.2009.00126.x
Show in context These are excreted by bacteria, plants and fungi, primarily to mobilise bioessential Fe (Haas, 2003) and to cope with metal toxicity (Braud et al., 2010).
View in article
Today, siderophores also enable microorganisms to reduce toxic metal accumulation and to increase metal tolerance (e.g., Braud et al., 2010), which might have been evolutionary advantageous also for early life.
View in article
Braun, J.-J., Pagel, M., Muller, J.-P., Bilong, P., Michard, A., Guillet, B. (1990) Cerium anomalies in lateritic profiles. Geochimica et Cosmochimica Acta 54, 781–795. https://doi.org/10.1016/0016-7037(90)90373-S
Show in context During pedogenesis and oxidative weathering, Ce(IV) may form insoluble cerianite (CeO2) which remains fixed in the regolith when the REY(III) are gradually mobilised (Braun et al., 1990), eventually producing Ce anomalies in normalised REY patterns of soil.
View in article
Byrne, J.M., Klueglein, N., Pearce, C., Rosso, K.M., Appel, E., Kappler, A. (2015) Redox cycling of Fe(II) and Fe(III) in magnetite by Fe-metabolizing bacteria. Science 347, 1473–1476. https://doi.org/10.1126/science.aaa4834
Show in context Biota facilitates a strong control on redox-sensitive elements, such as Fe (e.g., Byrne et al., 2015).
View in article
Christenson, E.A., Schijf, J. (2011) Stability of YREE complexes with the trihydroxamate siderophore desferrioxamine B at seawater ionic strength. Geochimica et Cosmochimica Acta 75, 7047–7062. https://doi.org/10.1016/j.gca.2011.09.022
Show in context A class of biogenic ligands that interacts strongly with oxide and silicate minerals (Brantley et al., 2001; Kraemer et al., 2014) and that solubilises Fe, REY and other high field strength elements (Yoshida et al., 2004; Christenson and Schijf, 2011; Bau et al., 2013; Kraemer et al., 2015, 2017) are the so-called “siderophores”.
View in article
Stability constants for REY-DFOB complexes are several orders of magnitude higher than those of mono- and dicarbonate REY complexes in seawater (Liu and Byrne, 1998), with log Kf of hexadentate complexes ranging from 10.09 for La to 15.19 for Lu at seawater ionic strength (Christenson and Schijf, 2011).
View in article
The stability constants of Y and Ho with DFOB are similar (Christenson and Schijf, 2011) and no significant fractionation would be expected, in contrast to our results. Yttrium, however, appears to have a higher initial accessibility during WRI than Ho (Bau et al., 1998), which could explain the Y fractionation during short-term WRI.
View in article
In contrast to the log Kf of REY(III)-DFOB (between 10.09 ± 0.08 and 15.19 ± 0.02; Christenson and Schijf, 2011) and Ce(III)-DFOB hexadentate complexes (log Kf ≈ 11.59; back-extrapolated from Nd(III)- and Pr(III)-DFOB data of Christenson and Schijf, 2011), log Kf of Ce(IV)-DFOB was estimated to be significantly higher (25–30; Yoshida et al., 2004) and thus comparable to Fe(III)-DFOB.
View in article
Duckworth, O.W., Sposito, G. (2005) Siderophore–Manganese (III) interactions. I. Air-Oxidation of Manganese(II) Promoted by Desferrioxamine B. Environmental Science and Technology 39, 6037–6044. https://doi.org/10.1021/es050275k
Show in context a lowering of the oxidation potential by DFOB-complexation and rapid oxidation by ambient air (“air-oxidation”; Duckworth and Sposito, 2005) which increases the metal-ligand complex stability (Hernlem et al., 1999; Kraemer et al., 2014), and.
View in article
Enzingmüller-Bleyl, T.C., Boden, J.S., Herrmann, A.J., Ebel, K.W., Sánchez-Baracaldo, P., Frankenberg-Dinkel, N., Gehringer, M.M. (2021) Lack of Fe(II) transporters in basal Cyanobacteria complicates iron uptake in ferruginous Archean oceans. (Preprint) bioRxiv, 2021.11.08.467730. https://doi.org/10.1101/2021.11.08.467730
Show in context Recent literature suggests that cyanobacteria may have produced siderophores for Fe(III) acquisition even before the Great Oxidation Event in the Palaeoproterozoic (Enzingmüller-Bleyl et al., 2021).
View in article
Farkas, E., Enyedy, E.A., Zékány, L., Deák, G. (2001) Interaction between iron(II) and hydroxamic acids: oxidation of iron(II) to iron(III) by desferrioxamine B under anaerobic conditions. Journal of Inorganic Biochemistry 83, 107–114. https://doi.org/10.1016/S0162-0134(00)00197-5
Show in context At the observed pH range, H4DFOB+ is the dominant species in solution, and anoxic Ce oxidation may follow a reaction path similar to the anoxic siderophore-promoted oxidation of ferrous Fe (Farkas et al., 2001).
View in article
German, C.R., Elderfield, H. (1990) Application of the Ce anomaly as a paleoredox indicator: The ground rules. Paleoceanography and Paleoclimatology 5, 823–833. https://doi.org/10.1029/PA005i005p00823
Show in context Due to their particle reactivity and the presumed immobility of REY, presence or absence of Ce anomalies in palaeoweathering profiles and marine chemical sediments are commonly used as a robust qualitative redox proxy (e.g., German and Elderfield, 1990; Riding et al., 2014).
View in article
Giese, U., Bau, M. (1994) Trace element accessibility in mid-ocean ridge and ocean island basalt: an experimental approach. Goldschmidt Abstracts 1994, 329–330. https://rruff.info/doclib/MinMag/Volume_58A/58A-1-329.pdf.
Show in context As REY are incompatible with respect to most rock-forming igneous minerals, they are, in rocks with porphyritic/phaneritic textures, not incorporated into the crystal lattice but are preferentially located along grain boundaries and in the interstitial crystal spaces (Giese and Bau, 1994).
View in article
In volcanic glass, however, Eu(II) and all REY(III) are homogenously distributed, and hence no EuCN anomaly can develop (Giese and Bau, 1994; Bach and Irber, 1998; Shibata et al., 2006; Kraemer et al., 2015).
View in article
Gledhill, M., van den Berg, C.M.G., Nolting, R.F., Timmermans, K.R. (1998) Variability in the speciation of iron in the northern North Sea. Marine Chemistry 59, 283–300. https://doi.org/10.1016/S0304-4203(97)00097-2
Show in context In modern seawater, complexation of bioessential micronutrients (e.g., Fe(III)) is dominated by organic ligands, and previous studies have suggested that this ligand pool is dominated by siderophores (van den Berg, 1995; Gledhill et al., 1998; Mawji et al., 2008).
View in article
Haas, H. (2003) Molecular genetics of fungal siderophore biosynthesis and uptake: the role of siderophores in iron uptake and storage. Applied Microbiology and Biotechnology 62, 316–330. https://doi.org/10.1007/s00253-003-1335-2
Show in context These are excreted by bacteria, plants and fungi, primarily to mobilise bioessential Fe (Haas, 2003) and to cope with metal toxicity (Braud et al., 2010).
View in article
Hernlem, B.J., Vane, L.M., Sayles, G.D. (1999) The application of siderophores for metal recovery and waste remediation: examination of correlations for prediction of metal affinities. Water Research 33, 951–960. https://doi.org/10.1016/S0043-1354(98)00293-0
Show in context a lowering of the oxidation potential by DFOB-complexation and rapid oxidation by ambient air (“air-oxidation”; Duckworth and Sposito, 2005) which increases the metal-ligand complex stability (Hernlem et al., 1999; Kraemer et al., 2014), and.
View in article
Kraemer, D., Kopf, S., Bau, M. (2015) Oxidative mobilization of cerium and uranium and enhanced release of “immobile” high field strength elements from igneous rocks in the presence of the biogenic siderophore desferrioxamine B. Geochimica et Cosmochimica Acta 165, 263–279. https://doi.org/10.1016/j.gca.2015.05.046
Show in context A class of biogenic ligands that interacts strongly with oxide and silicate minerals (Brantley et al., 2001; Kraemer et al., 2014) and that solubilises Fe, REY and other high field strength elements (Yoshida et al., 2004; Christenson and Schijf, 2011; Bau et al., 2013; Kraemer et al., 2015, 2017) are the so-called “siderophores”.
View in article
Siderophore-promoted oxidation of Ce under oxic conditions has been described in literature (Brantley et al., 2001; Yoshida et al., 2004; Bau et al., 2013; Kraemer et al., 2015, 2017).
View in article
patterns of the anoxic to oxic experiments at the same time interval are similar, except for Ce, and corroborate published data from oxic experiments (Kraemer et al., 2015).
View in article
In the course of individual experiments, pH first decreased from 5.5 to <5, then increased and mostly remained in the range of 5–7 after 24 hr (Figs. S-2–S-4, Table S-4), similar to previous studies (Kraemer et al., 2015).
View in article
In volcanic glass, however, Eu(II) and all REY(III) are homogenously distributed, and hence no EuCN anomaly can develop (Giese and Bau, 1994; Bach and Irber, 1998; Shibata et al., 2006; Kraemer et al., 2015).
View in article
Siderophore-mediated oxidation was also demonstrated during oxic WRI with igneous rocks (Bau et al., 2013; Kraemer et al., 2015) and during scavenging onto Mn (hydr)oxides (Kraemer et al., 2017) with the development of a positive CeN anomaly.
View in article
a disequilibrium between Ce3+aq and Ce4+aq at the mineral–water interface, where minute amounts of tetravalent Ce in solution are strongly complexed by DFOB, facilitating further disequilibrium and Ce4+ regeneration by continuous oxidation of Ce3+ to maintain redox equilibrium (“siderophore redox pump”; Bau et al., 2013; Kraemer et al., 2015, 2017).
View in article
The only distinct feature, apart from the Ce anomaly, is the occasionally very strong depletion of La (Bau et al., 2013; Kraemer et al., 2015, 2017; this study).
View in article
Kraemer, D., Tepe, N., Pourret, O., Bau, M. (2017) Negative cerium anomalies in manganese (hydr)oxide precipitates due to cerium oxidation in the presence of dissolved siderophores. Geochimica et Cosmochimica Acta 196, 197–208. https://doi.org/10.1016/j.gca.2016.09.018
Show in context A class of biogenic ligands that interacts strongly with oxide and silicate minerals (Brantley et al., 2001; Kraemer et al., 2014) and that solubilises Fe, REY and other high field strength elements (Yoshida et al., 2004; Christenson and Schijf, 2011; Bau et al., 2013; Kraemer et al., 2015, 2017) are the so-called “siderophores”.
View in article
Siderophore-promoted oxidation of Ce under oxic conditions has been described in literature (Brantley et al., 2001; Yoshida et al., 2004; Bau et al., 2013; Kraemer et al., 2015, 2017).
View in article
Siderophore-mediated oxidation was also demonstrated during oxic WRI with igneous rocks (Bau et al., 2013; Kraemer et al., 2015) and during scavenging onto Mn (hydr)oxides (Kraemer et al., 2017) with the development of a positive CeN anomaly.
View in article
a disequilibrium between Ce3+aq and Ce4+aq at the mineral–water interface, where minute amounts of tetravalent Ce in solution are strongly complexed by DFOB, facilitating further disequilibrium and Ce4+ regeneration by continuous oxidation of Ce3+ to maintain redox equilibrium (“siderophore redox pump”; Bau et al., 2013; Kraemer et al., 2015, 2017).
View in article
The only distinct feature, apart from the Ce anomaly, is the occasionally very strong depletion of La (Bau et al., 2013; Kraemer et al., 2015, 2017; this study).
View in article
Kraemer, S.M., Duckworth, O.W., Harrington, J.M., Schenkeveld, W.D.C. (2014) Metallophores and Trace Metal Biogeochemistry. Aquatic Geochemistry 21, 159–195. https://doi.org/10.1007/s10498-014-9246-7
Show in context A class of biogenic ligands that interacts strongly with oxide and silicate minerals (Brantley et al., 2001; Kraemer et al., 2014) and that solubilises Fe, REY and other high field strength elements (Yoshida et al., 2004; Christenson and Schijf, 2011; Bau et al., 2013; Kraemer et al., 2015, 2017) are the so-called “siderophores”.
View in article
a lowering of the oxidation potential by DFOB-complexation and rapid oxidation by ambient air (“air-oxidation”; Duckworth and Sposito, 2005) which increases the metal-ligand complex stability (Hernlem et al., 1999; Kraemer et al., 2014), and.
View in article
Liu, X., Byrne, R.H. (1998) Comprehensive Investigation of Yttrium and Rare Earth Element Complexation by Carbonate Ions Using ICP–Mass Spectrometry. Journal of Solution Chemistry 27, 803–815. https://doi.org/10.1023/A:1022677119835
Show in context Stability constants for REY-DFOB complexes are several orders of magnitude higher than those of mono- and dicarbonate REY complexes in seawater (Liu and Byrne, 1998), with log Kf of hexadentate complexes ranging from 10.09 for La to 15.19 for Lu at seawater ionic strength (Christenson and Schijf, 2011).
View in article
Martell, A., Smith, R. (2001) NIST Stability Constants of Metal Complexes Database 46, v. 6.0. National Institute of Standards and Technology, Gaithersburg, MD.
Show in context In comparison, the hexadentate coordination of Fe3+ results in a stability constant of 30.6 (DFOB; Martell and Smith, 2001).
View in article
Mawji, E., Gledhill, M., Milton, J.A., Tarran, G.A., Ussher, S., Thompson, A., Wolff, G.A., Worsfold, P.J., Achterberg, E.P. (2008) Hydroxamate Siderophores: Occurrence and Importance in the Atlantic Ocean. Environmental Science & Technology 42, 8675–8680. https://doi.org/10.1021/es801884r
Show in context In modern seawater, complexation of bioessential micronutrients (e.g., Fe(III)) is dominated by organic ligands, and previous studies have suggested that this ligand pool is dominated by siderophores (van den Berg, 1995; Gledhill et al., 1998; Mawji et al., 2008).
View in article
Neaman, A., Chorover, J., Brantley, S.L. (2005) Implications of the evolution of organic acid moieties for basalt weathering over geological time. American Journal of Science 305, 147–185. https://doi.org/10.2475/ajs.305.2.147
Show in context Neaman et al. (2005) argued that organisms have had to cope with low nutrient bioavailability since at least the colonisation of the land surface.
View in article
Nelson, M., Cooper, C.R., Crowley, D.E., Reid, C.P.P., Szaniszlo, P.J. (1988) An escherichia coli bioassay of individual siderophores in soil. Journal of Plant Nutrition 11, 915–924. https://doi.org/10.1080/01904168809363856
Show in context In (modern) soils, siderophores are at least as abundant as in marine systems, with concentrations up to the mmol/L range (Nelson et al., 1988), and significantly enhance the mobility of trace elements such as REY (Wiche et al., 2017).
View in article
Riding, R., Fralick, P., Liang, L. (2014) Identification of an Archean marine oxygen oasis. Precambrian Research 251, 232–237. https://doi.org/10.1016/j.precamres.2014.06.017
Show in context Due to their particle reactivity and the presumed immobility of REY, presence or absence of Ce anomalies in palaeoweathering profiles and marine chemical sediments are commonly used as a robust qualitative redox proxy (e.g., German and Elderfield, 1990; Riding et al., 2014).
View in article
Shibata, S.-N., Tanaka, T., Yamamoto, K. (2006) Crystal structure control of the dissolution of rare earth elements in water-mineral interactions. Geochemical Journal 40, 437–446. https://doi.org/10.2343/geochemj.40.437
Show in context In volcanic glass, however, Eu(II) and all REY(III) are homogenously distributed, and hence no EuCN anomaly can develop (Giese and Bau, 1994; Bach and Irber, 1998; Shibata et al., 2006; Kraemer et al., 2015).
View in article
van den Berg, C.M.G. (1995) Evidence for organic complexation of iron in seawater. Marine Chemistry 50, 139–157. https://doi.org/10.1016/0304-4203(95)00032-M
Show in context In modern seawater, complexation of bioessential micronutrients (e.g., Fe(III)) is dominated by organic ligands, and previous studies have suggested that this ligand pool is dominated by siderophores (van den Berg, 1995; Gledhill et al., 1998; Mawji et al., 2008).
View in article
Wang, X., Barrat, J.-A., Bayon, G., Chauvaud, L., Feng, D. (2020) Lanthanum anomalies as fingerprints of methanotrophy. Geochemical Perspectives Letters 14, 26–30. https://doi.org/10.7185/geochemlet.2019
Show in context Wang et al. (2020) identified La anomalies as fingerprints for methanotrophic activity.
View in article
Wiche, O., Tischler, D., Fauser, C., Lodemann, J., Heilmeier, H. (2017) Effects of citric acid and the siderophore desferrioxamine B (DFO-B) on the mobility of germanium and rare earth elements in soil and uptake in Phalaris arundinacea . International Journal of Phytoremediation 19, 746–754. https://doi.org/10.1080/15226514.2017.1284752
Show in context In (modern) soils, siderophores are at least as abundant as in marine systems, with concentrations up to the mmol/L range (Nelson et al., 1988), and significantly enhance the mobility of trace elements such as REY (Wiche et al., 2017).
View in article
Yoshida, T., Ozaki, T., Ohnuki, T., Francis, A.J. (2004) Adsorption of rare earth elements by γ-Al2O3 and Pseudomonas fluorescens cells in the presence of desferrioxamine B: implication of siderophores for the Ce anomaly. Chemical Geology 212, 239–246. https://doi.org/10.1016/j.chemgeo.2004.08.046
Show in context A class of biogenic ligands that interacts strongly with oxide and silicate minerals (Brantley et al., 2001; Kraemer et al., 2014) and that solubilises Fe, REY and other high field strength elements (Yoshida et al., 2004; Christenson and Schijf, 2011; Bau et al., 2013; Kraemer et al., 2015, 2017) are the so-called “siderophores”.
View in article
Siderophore-promoted oxidation of Ce under oxic conditions has been described in literature (Brantley et al., 2001; Yoshida et al., 2004; Bau et al., 2013; Kraemer et al., 2015, 2017).
View in article
Yoshida et al. (2004) conducted adsorption experiments of REY-DFOB complexes on γ-Al2O3 and P. fluorescens and found that Ce behaves markedly different from La(III) and Pr(III).
View in article
In contrast to the log Kf of REY(III)-DFOB (between 10.09 ± 0.08 and 15.19 ± 0.02; Christenson and Schijf, 2011) and Ce(III)-DFOB hexadentate complexes (log Kf ≈ 11.59; back-extrapolated from Nd(III)- and Pr(III)-DFOB data of Christenson and Schijf, 2011), log Kf of Ce(IV)-DFOB was estimated to be significantly higher (25–30; Yoshida et al., 2004) and thus comparable to Fe(III)-DFOB.
View in article
top
Supplementary Information
The Supplementary Information includes:
Download Table S-4 (Excel).
Download the Supplementary Information (PDF).
Figures
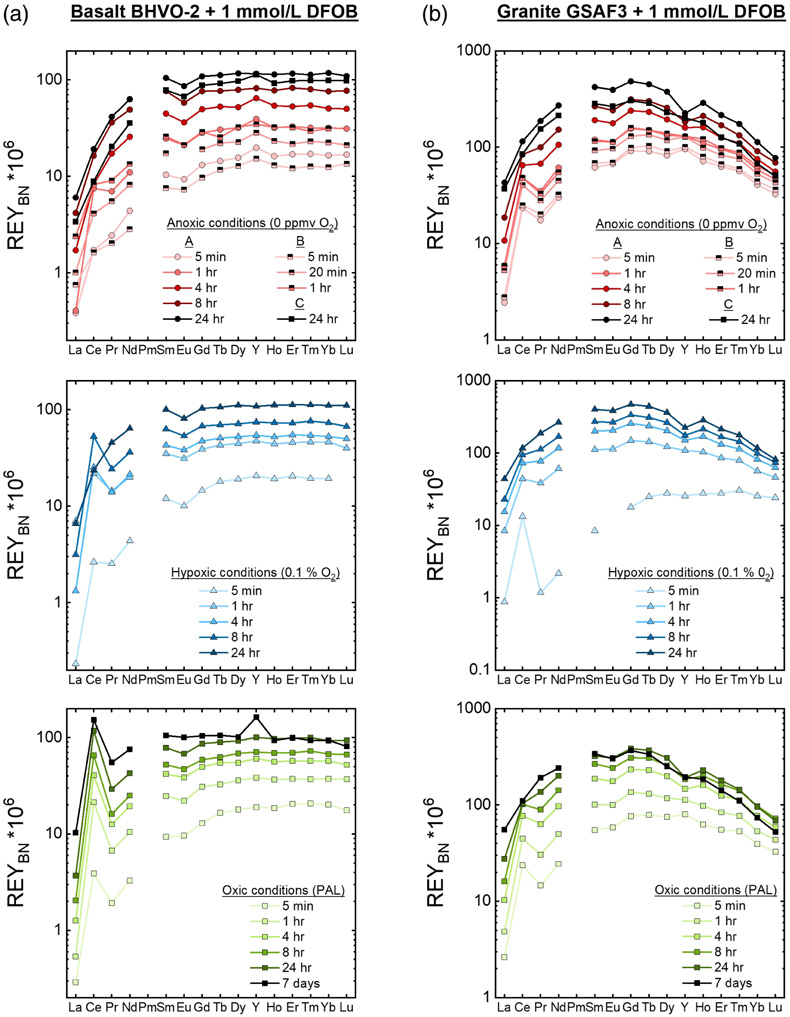
Figure 1 REYBN patterns of time series experiments with (a) basalt and (b) granite.
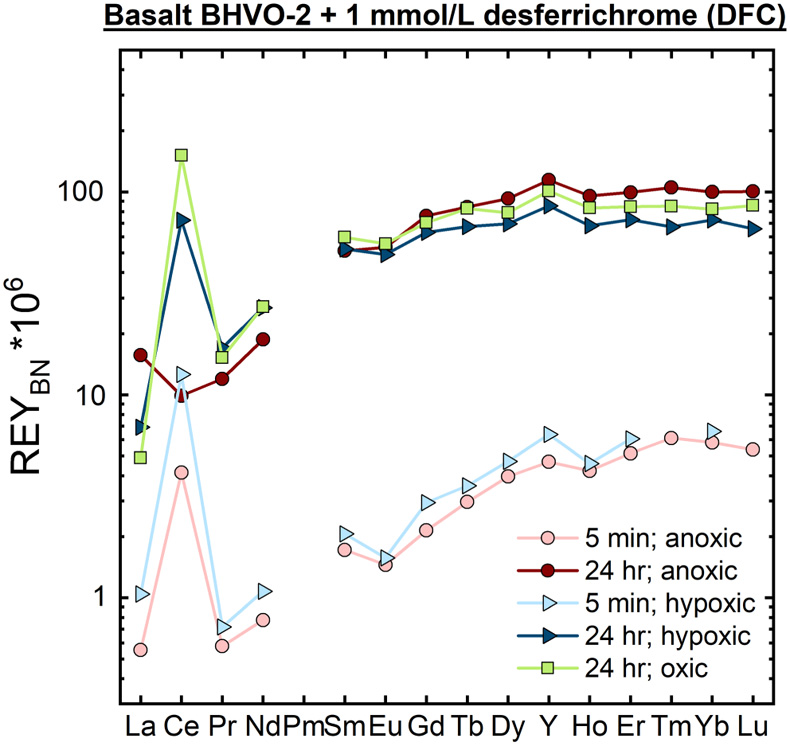
Figure 2 REYBN patterns of experiments with basalt and DFC.
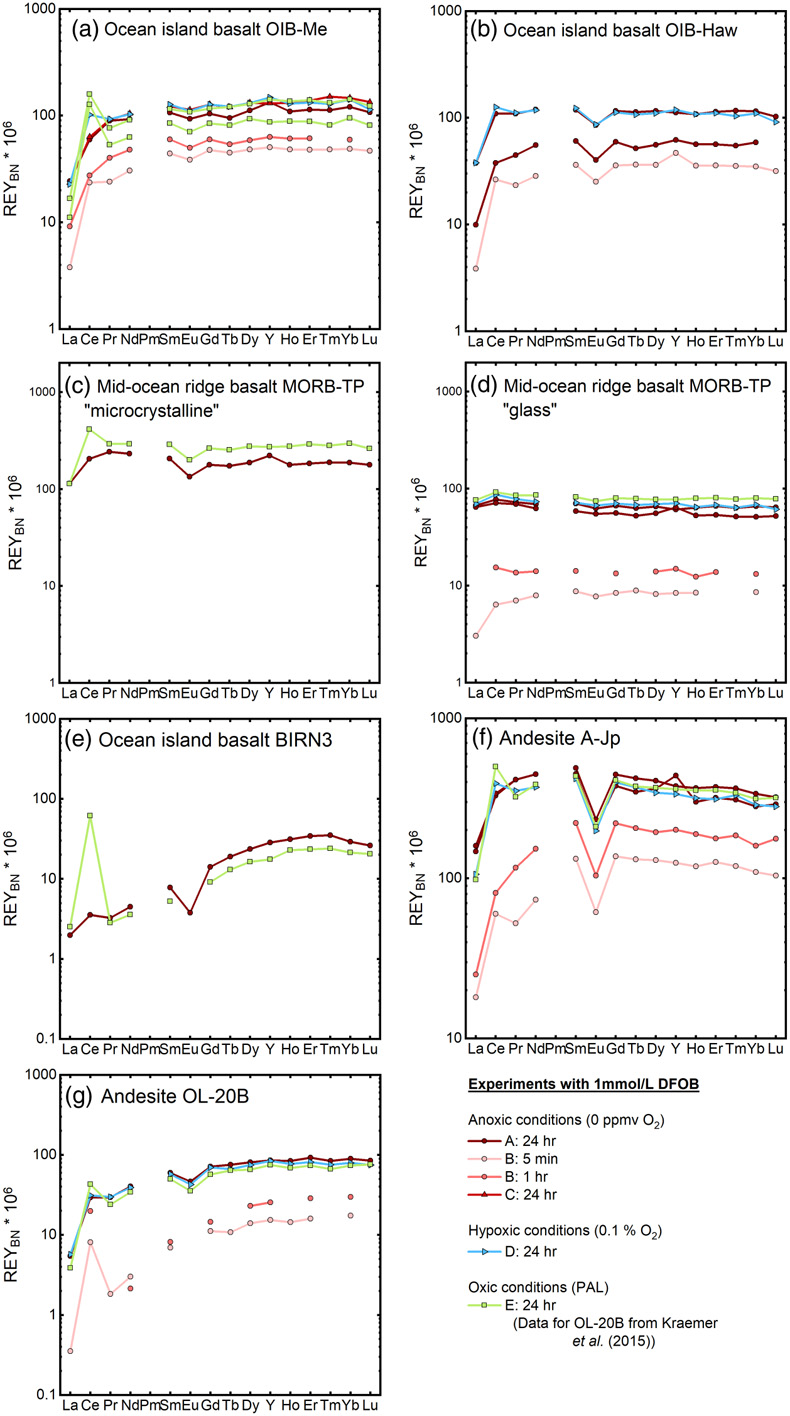
Figure 3 REYBN patterns of leaching experiments with different igneous rocks. The letters A-E in the legend refer to different experiment runs. Details are listed in Table S-2.
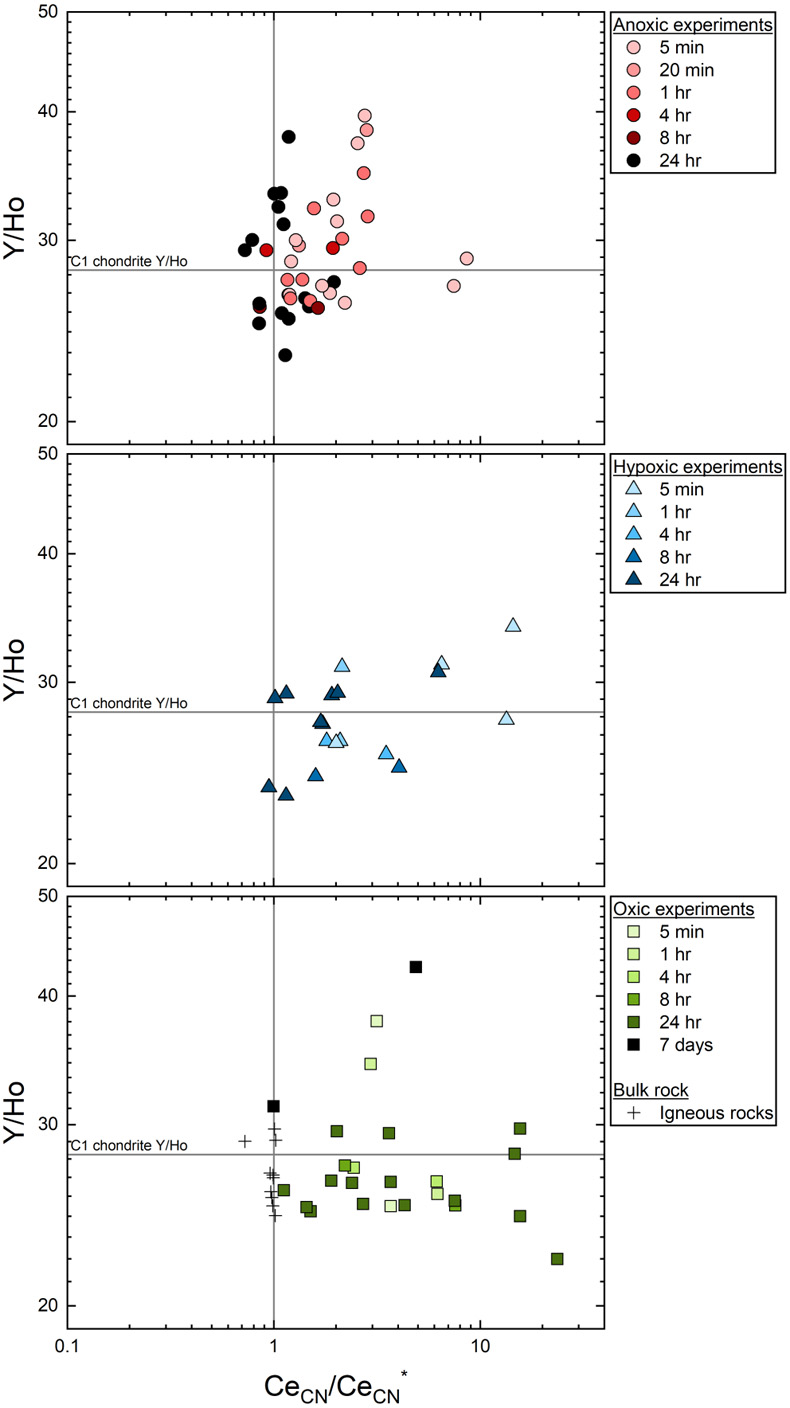
Figure 4 Graph of CeCN/CeCN* vs. Y/Ho ratio for bulk rocks and leachates.